1. Introduction
Granules are formed by the interaction of microbial cells of different or similar strains through basic mechanism of autoaggregation (AAg) or coaggregation. AAg refers to the physical cell-to-cell interaction between genetically identical cells, whereas coaggregation refers to the interaction between genetically distinct bacterial cells. Microbial aggregation is a part of initial cell interaction which results in granule formation. Several factors such as type of seed sludge, type and concentration of substrate, presence of extracellular polymeric substrate (EPS), composition of the media, pH, temperature and operational set-up of the reactor can affect the initial cell interactions. Additionally, surface hydrophobicity (SHb) can also influence granule formation. Cell aggregation and SHb reportedly have a good correlation to the adhesion capacity of bacteria [1–3]. The factors that influence the adhesion ability of microbials cells have been widely investigated and could be measured through microbial aggregation and SHb [2, 4–10].
Granular sludge formed by cell immobilization consist of biofilm, entrapped microorganisms and microbial aggregates [11]. However, it differs from biofilm formation due to the absence of carrier materials [5, 12–15]. EPS is one of the biofilm components found in granular sludge [8, 16–17]. EPS can cause cell SHb and act as a driving force during biogranulation [5, 8–9, 18–19]. Many studies have reported that EPS and SHb are correlated. They facilitate the aggregation of bacteria and maintenance of the granular structure [8, 18, 20–21]. Several efforts have been made to enhance granulation with additives and change the start up or operating conditions either in aerobic or anaerobic systems [12]. Some of the notable additives are cations [22–26] and synthetic polymers [27–29]. Cations can facilitate the bridging of the negatively charged groups on the cell surface with the secreted EPS [23, 30–31]. This enables the initial cell interaction stage of granulation process. Nomura et al. 2009 reported that the aggregation of washed cells (Lactococcus lactis JCM 5805) slightly increased at high concentration of cations. In contrast, the aggregation of microbial cell in unwashed cell decreased with increasing cationic strength. Additionally, when EPS was present in suspensions of unwashed cells, a low concentration of cations were found during the aggregation of the microbial cells due to an attractive bridging force. From the application viewpoint, this technology could be useful in flocculation of dispersed sludge, and binding of negatively charged cells to promote the granulation process using cations (Ca2+, Mg2+, Fe2+ and Fe3+). It is also applicable in bacterial adhesion in wastewater treatment [32].
Granular formation require a long development time depending on the experimental conditions and set-up. In some cases, granules are formed in weeks or months, but require sufficient time to grow and increase in size. Previous studies have reported that bacterial behavior or characteristic is independent of the changes in the environment [33–34].
The aim of this study was to investigate the reaction of a few selected bacteria to any change or modifications in its environment. The effect of various cations (Ca2+, Mg2+, Al3+, Mn2+ and Zn2+) on microbial aggregation and SHb of three bacteria (Brevibacillus panacihumi strain (ZB1), Lysinibacillus fusiformis strain (ZB2) and Enterococcus faecalis strain (ZL)) were examined in order to improve the aerobic granulation start-up. Statistical experimental design was used to provide a quantitative understanding of the main interaction effects of the input variables and to develop the correlation between the input variables and responses.
2. Materials and Methods
2.2. Preparation of Bacterial Inoculum
The microorganisms used in this study were ZB1, ZB2 and ZL. Each bacteria was grown overnight in nutrient broth. The cultures were agitated in a rotary shaker at room temperature until the exponential growth phase was reached.
2.3. Reactor Set-up
The experiment was performed using a 1 L schott bottle as reactor. Oxygen was supplied to the reactor at a fixed aeration rate of 2 L/min, in a manner similar to that of Sequential batch reactor (SBR) system in order to mimick the SBR granulation technique and process. The working volume of the experiment was 250 mL (10%v/v of inoculum was used). The bacteria was incubated until the stationary phase was attained, thereafter, analysis was carried out.
2.4. Divalent and Trivalent Cation Solutions
The effect of divalent and trivalent cations on microbial AAg and SHb of three bacteria were investigated using aluminum chloride (AlCl3), calcium chloride (CaCl2), magnesium sulfate (MgSO4), manganese chloride (MnCl2) and zinc chloride (ZnCl2). A stock solution of 100 g/L was prepared for each cation and the working concentrations were prepared daily by means of dilution. The experimental conditions for cationic concentration and SHb during the initial cell interaction of the biogranulation pocess was based on previous study [25, 31, 36–37]. All cation solutions were sterilized at 121°C for 5 min [36] as shown in Table 1.
2.5. Aggregation Test
Aggregation tests were conducted to represent the adhesion ability of bacteria under the influence of divalent and trivalent cations based on the procedure used by Chen et al. [1] and Rahman et al. [2]. Each culture was incubated in the bioreactor at an aeration rate of 2 L/min. From each sample, about 10 mL of the culture was taken from the bioreactor and centrifuged at a slow centrifugation speed of 650 g for 2 min as described by Malik et al. [38]. Turbidity measurement was used as an indcator of the optical density (OD) of each sample at the initial and final stationary phase of bacteria growth.
The aggregation ability was expressed as AAg percentage (AAg%) and was calculated using Eq. (1):
Where
AAg% is the percentage of aggregation,
A0 is the absorbance of cultured media at 0 h,
A is the absorbance of cultured media after centrifugation
For the AAg%, the culture can be classified into three groups: high coaggregation (HAg: > 70% Ag), medium aggregation (MAg: 20–70% Ag) and low aggregation (LAg: < 20% Ag) cultures. A high aggregation index denotes a strong tendency of the cells to agglomerate into an aggregate [7].
2.6. SHb Test
The SHb of the bacterial strains were based on the microbial adhesion to hydrocarbon assay. The SHb of bacteria was determined according to the methods described by Rahman et al. [2] and was modified to suit the sample used in this study.
Each bacteria culture was incubated in the aerated bioreactor. 10 mL of each sample was taken from the glass bottle (bioreactor) after the growth of bacteria reached the stationary phase. The bacterial cells were harvested by centrifugation at 4,000 rpm for 10 min. The pellets obtained were washed twice with 50 mM of K2HPO4 (pH 7.0) and then resuspended in the same buffer to obtain an absorbance of about 0.5 at 660 nm. Five (5) mL of bacterial suspension was mixed with 1 mL of xylene (C6H4(CH3)2) by vortexing for 120 s and then allowed to stand for 1 h at room temperature. The absorbance of the bacterial suspension in the aqueous phase after mixing (S) was compared to the absorbance taken at the initial stage of the experiment (S0). Changes in absorbance due to the bacterial adhesion to the hydrocarbons was measured at 660 nm by using an OD Meter (DR5000 HACH Spectrophometer). All the samples were analysed in duplicates. The SHb was expressed as SHb% and calculated by Eq (2):
Where
2.7. Experimental Procedure
All three bacteria were cultured separately in nutrient broth until the OD was close to 1. Thereafter, about 25 mL (10% v/v) of the cultured sample was inoculated in a separate 250 mL Schott bottle containing synthetic wastewater. The inoculated synthetic sample was allowed to incubate under aerobic condition until it reached the stationary phase. The aggregation and SHb assay were then carried out. Instead of using physical shaking as commonly reported in previous aggregation test [2, 10, 39–40], aeration was used to provide a close resemblance to the techniques used in typical granulation process. 10 mL of each sample was withdrawn after the stationary phase was reached for the aggregation and SHb tests. To determine the effect of Ca2+, Mg2+, Al3+, Mn2+ and Zn2+ on aggregation and SHb of ZL, ZB1 and ZB2 cultures, an experimental design was employed using Minitab® (Version 16.3.2). This approach showed the effect of each factor or input variable separately (termed as the main effect) and the effect of the interaction between the variables (termed as the interaction effect) on the responses (i.e., aggregation and SHb). Thus, the outcome was quantitatively determined. The experimental design followed a 2-level factorial design (coded as −1 and +1) comprising of 64 runs. The Ca2+, Mg2+, Al3+, Mn2+ and Zn2+ range of values used in the experiments are shown in Table 2. The responses were measured as percentage aggregation (%Ag) and percentage SHb (%SHb).
3. Results and Discussion
3.1. AAg and SHb of ZL, ZB1 and ZB2 Strains in Synthetic Wastewater
ZL, ZB1 and ZB2 were obtained from the decolourising bacteria isolated from textile wastewater and palm oil mill effluent [41–43].
3.1.1. Growth profile of ZL, ZB1 and ZB2 in synthetic wastewater
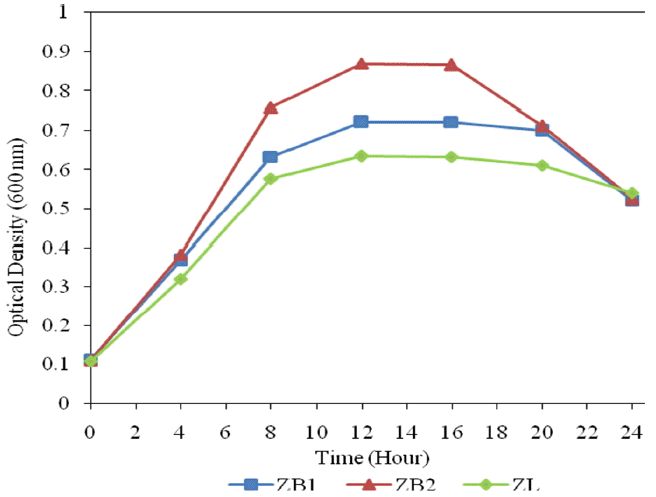
Each bacteria was grown in synthetic wastewater with the addition of 10% (v/v) nutrient broth under aerobic condition for a 24 h period. Individual bacterial growth was measured using spectrophotometer at a wavelength of 600 nm. The growth profiles were determined by plotting the value of OD600 vs time. The growth profile of the various bacteria (ZL, ZB1 and ZB2) were obtained as shown in Fig. 1. The bacteria reached the stationary phase in 12 h.
The AAg and SHb of the strains ZB1, ZB2 and ZL are shown in Table 3. These bacteria can be classified under the medium rank bacteria aggregation.
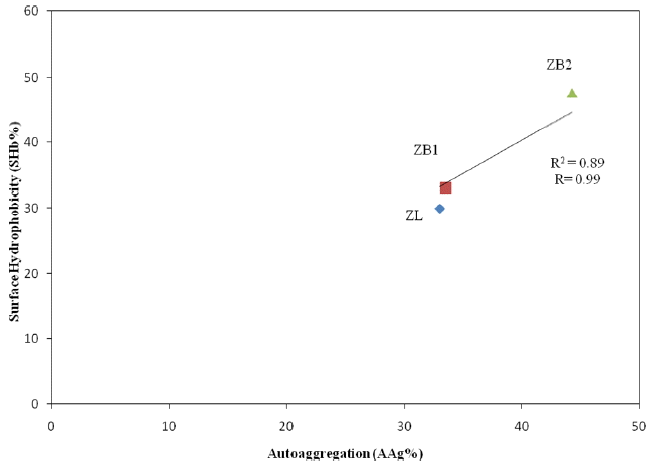
The plot of the measured AAg% vs the SHb% of the various strains indicates the extent of microbial adhesion to hydrocarbons. Correlation between AAg and SHb for each strain (ZB1, ZB2 and ZL) was also calculated. A correlation (r2) of 0.89 was obtained between AAg and SHb for the various strains. This result indicates that a good correlation exist between AAg and SHb. Thus, SHb could be one of the determinants of AAg. The result obtained is in agreement with that of Rahman et al. [2].
Fig. 2 and Table 3 show the interdependece of AAg% and SHb% on each other. Thus, SHb% can be referred to AAg% for the various bacteria investigated. However, this hypothesis does not prove that it is applicable to other bacteria. SHb of bacteria can also be caused by the presence of EPS which notably causes adhesion between bacteria and floc [9]. Current understanding is based on a polymer-bridging model, where microbial aggregation is interpreted from the result of interactions of naturally produced EPS and microbial cells. EPS excreted by microbial cells play a key role in adsorbing and bridging cell surfaces. A direct correlation exist between EPS accumulation and microbial aggregation as well as bacteria SHb.
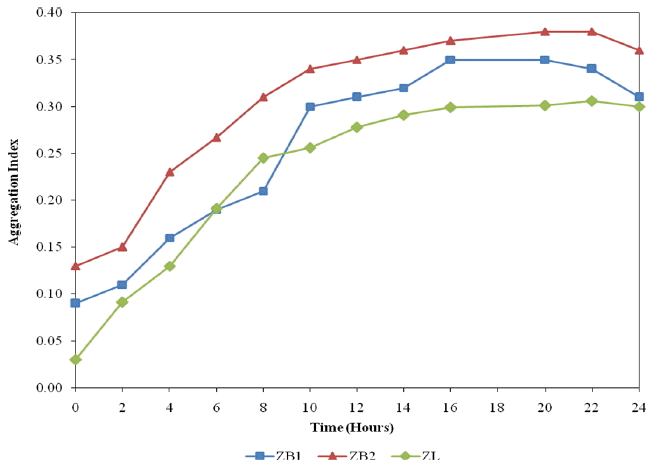
Fig. 3 shows the changes in AAg% with time for ZB1, ZB2 and ZL cultures under aerobic conditions. AAg% in ZB1, ZB2 and ZL bacteria showed an increasing trend with time at the exponential growth phase prior to the stationary growth phase. During the stationary growth phase, there was not much increase in the AAg%. A similar observation was made by Chen et al. [1] who noted that the autoaggregation of Aeromonas punctata strain T394 and Ochrobactrum anthropi strain WZR occurred in the activated sludge system.
Based on the categories [2], ZL, ZB1 and ZB2 were found in the medium AAg group. Although the various bacteria fall in the same rank, AAg% for each bacteria was different. The sequence of AAg% for the three bacteria were ZB2 > ZB1 > ZL. ZB2 also ranked highest during the incubation.
The AAg of the various bacteria could further be compared through their morphology as shown in Fig. 4. Fig. 4(a) – Fig. 4(c) represent the morphological structure of ZL, ZB1 and ZB2, respectively. The results show that the lowest AAg% occurred in ZL due to small number of flocs whereas ZB1 had a cell aggregate formation with irregular-shaped flocs similar to ZB2. However, ZB2 appeared to have a dense clump of cells. Evidently, ZB2 had the most cell aggregate formation with irregular-shaped floc.
3.2. Factorial Design Analysis
3.2.1. Effect of cations on AAg and SHb of ZL, ZB1 and ZB2 bacteria
Previous studies utilized physical shaking to generate shear force [2, 7, 38]. In order to mimic the real situation of the aerobic granulation process, physical shaking method was replaced with aeration in this study. This was to ensure consistent results comparable to conventional methods. Aeration was used to introduce the shear effect onto the microorganisms in the bioreactor. Furthermore, it helped to increase the contact angle of collision between microorganisms and cations which accelerated the aggregation process.
The effect of the cations on these variables (AAg and SHb) were presented by the responses produced. When the effect of one input variable affects the responses, there is an obvious interaction effect between the input variables. The results from the factorial design were analyzed by the analysis of variance (ANOVA) to provide information on the estimated effects with their corresponding coefficients. ANOVA gave a summary of the significance of the main and interaction effects measured by the P-value. The responses obtained were statistically evaluated at 90% confidence level (P-value < 0.1). Table 4 shows the experimental result of the factorial design of the study. Each of the independent variable was investigated at a high (+1) and low (−1) level.
3.2.2. The main effect of cations on AAg of ZL, ZB1 and ZB2
Table 5 shows the five main effects of cations on AAg for ZL, ZB1 and ZB2 (ANOVA summary).
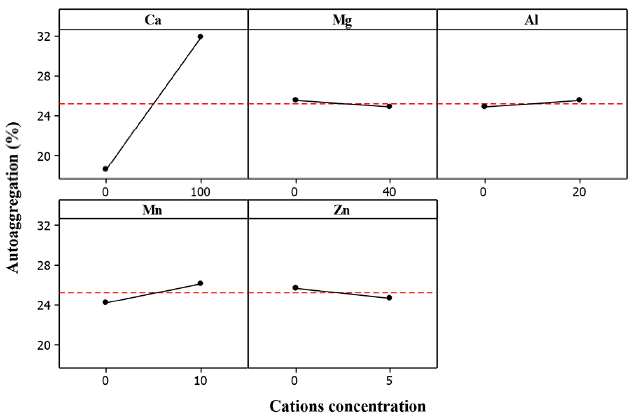
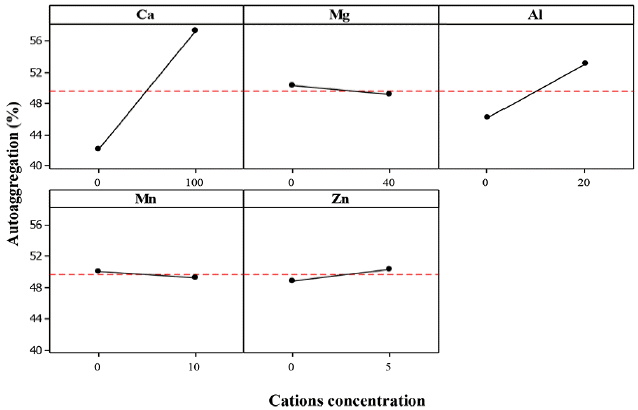
Calcium cation (Ca2+) showed a significant effect with P-values less than 0.10 in the ZL, ZB1 and ZB2 cultures. Thus, an increase of Ca2+ concentration increased the AAg%. It is well known that Ca2+ is an important cation for the aggregation process and can increase the bioflocculation of most bacteria [25, 31, 44]. Mckinney and Horwood [45] and Tezuka et al. [44] proposed the rule of divalent cationic bridging function (DCB theory) which involves the interaction between divalent cations and negatively charged functional group of the EPS. In most biogranulation development; Ca2+ promotes the formation of cell-to-cell bridging, improve aggregation and stabilization of the biopolymer, enhance bioflocculation and the overall granulation process [30, 44–46].
The effect of magnesium cation (Mg2+) on the AAg was not significant in each of the experimented bacteria. This result is in contrast with that of Li et al. [47], which stated that Mg2+ had similar effect as Ca2+ on microbial aggregation which helps in the cell-to-cell bridging of microorganisms. Additionally, it has been reported that Mg2+ accelerated the formation of aerobic granules [23]. In this study, Mg2+ had a weak effect on the structure of granules and could not play a bridging and core-induced role. This may imply that magnesium enhanced the sludge granulation process through biochemical function. Although Ca2+ and Mg2+ cations have similar properties, their respective interaction with biopolymers or EPS differ [48]. The bound water content of sludge was not affected by Mg2+ addition, but was remarkably reduced with Ca2+ was addition [23]. Although the effect of Mg2+ on AAg in the various bacteria was not statistically significant, Mg2+ is an important trace element and a source of nutrient for metabolic enzymes during the growth of microorganisms [15, 23].
The effect of aluminum cation (Al3+) on the AAg of ZB1 and ZB2 was significant, but was non-significant for ZL. The estimated main effects for ZB1 and ZB2 were 6.921 and 3.891, respectively. Fig. 5–7 show that the AAg of ZB1 and ZB2 increased with increasing Al3+. It has been reported that Al3+ can be used to reduce the required time for granule formation and increase aggregation rate [49]. Unlike most metal cations, Al3+ is not required either in the macro or even micro-organisms as nutrients in biological growth [50].
The effect of manganese cation (Mn2+) on the AAg of ZL culture was significant, but non-significant for ZB1 and ZB2 (Fig. 5). Mn2+ is among the essential nutrient required for microbial growth [50]. Huang et al. [51] utilized Mn2+ for augmentation during aerobic granule development. The authors found that Mn2+ augmentation could enhance the biogranulation process by stimulating the microbial diversity such as cell bridging and EPS secretion. Although there was a slight increase in AAg%, Mn2+ demonstrated the ability to cause microbial aggregation in the ZL culture. However the effect was largely different with that of Ca2+.
The effect of Zinc cation (Zn2+) on the AAg of the various bacteria was not significant. This could be attributed to inappropriate concentration of Zn2+ which can inhibit microorganisms or bacteria activity. In addition, excess concentration of Zn2+ resulted to a decrease in EPS production and loosening of aggregated cells [52].
3.2.3. The main effect of cations on SHb of ZL, ZB1 and ZB2
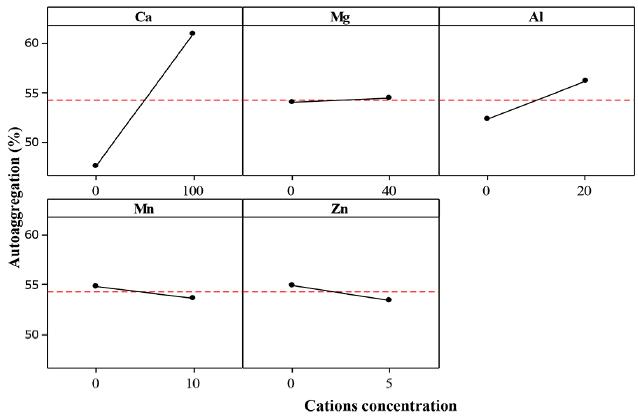
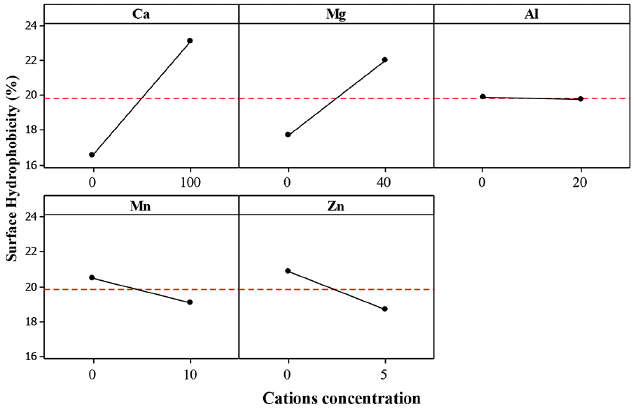
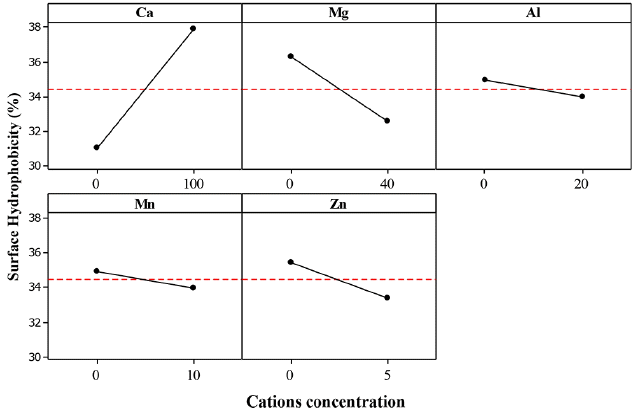
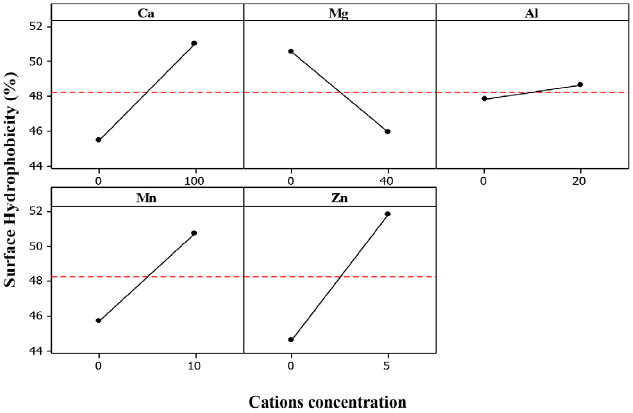
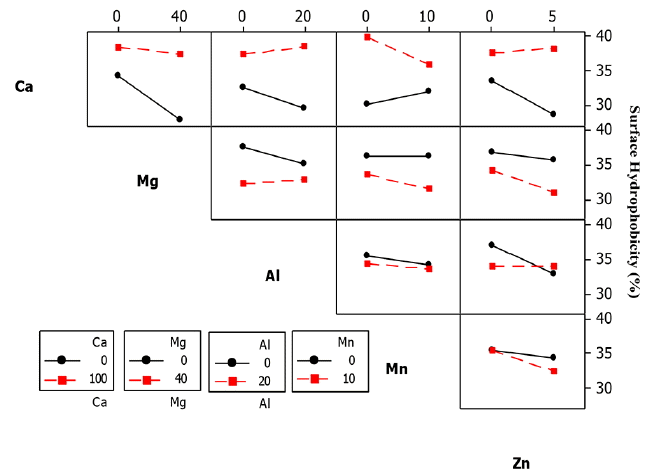
The effect of calcium cations (Ca2+) on the SHb for the three bacteria (ZL, ZB1 and ZB2) was significant with estimated main effect of 6.556, 6.882 and 5.515, respectively (Table 6).
From the statistical analysis, it could be explained that addition of Ca2+ caused an increase in Zl, ZB1 and ZB2 SHb% (Fig. 8–10).
Ca2+ cations acted as a binder between EPS and cells by increasing the cell aggregation. At higher bacteria aggregation, higher hydrophobicity occurs which strengthens cell to cell interaction within the microbial structure [9, 35]. It serves as a protective mechanism for the cells against unfavorable environmental conditions. Higgins and Novak [53] reported that the addition of Ca2+ to activated sludge caused a decrease of the bound water content which increased aggregation.
It is well known that the presence of EPS can enhance cell SHb [18]. EPS is produced when the bacteria is under stress conditions. Although Ca2+ does not produce EPS, the increase in Ca2+ may create stress condition on bacteria. This could increase the production of the EPS due to the changing environmental conditions [54]. The three bacteria strains used in this study could have been exposed to stress condition due to the presence of excess Ca2+. This was evident by the production of EPS and the subsequent SHb occurrence. Fig. 10 shows that the bonding between EPS and the aggregate cells increased with increasing Ca2+ concentration, suggesting that Ca2+ may create a stronger bond.
The result of the factorial design for the effect of calcium (Ca2+) on SHb was significant for all three bacteria experimented. The cell- to-cell bridging with the negatively charged sites on the EPS and bacteria surfaces increased with increasing Ca2+ concentration [51, 55]. Additionally, Ca2+ served as a cofactor or enzyme to induce the bacterial growth. Li et al. [48] reported that high EPS level was detected in anaerobic and aerobic granulation process in the presence of excess Ca2+ because the carbon utilization shifted towards EPS production. Increase in the cell-to-cell bridging between EPS and bacteria caused an increase in the SHb and the bacteria became more hydrophobic which facilitated adhesion or aggregation process [9]. The effect of Ca2+ had the same pattern with regards to the responses (AAg% and SHb%) in the presence of EPS. It is well known that EPS can capably mediate both the cohesion and adhesion of cells and play a fundamental role in sustaining the structural integrity in the development of biofilm, anaerobic and aerobic granules [38, 56].
Table 6 shows that the effect of Mg2+ on the SHb of ZL and ZB1 was significant, but non-significant for ZB2. The estimated main effect for ZL and ZB1 were 4.338 and 3.725, respectively. From the statistical analysis, addition of Mg2+ did not increase the SHb% of ZB2. Although the estimated effect of Mg2+ on SHb of ZB1 was negative, the effect was significant because it probably yielded a good interaction with other cations (Fig. 9). The Mg2+ cations promoted microbial aggregation and acted as a bridge for other bacteria surfaces and EPS. Additionally, the presence of Mg2+ produced higher amounts of EPS. However, further increase of Mg2+ did not yield an increase in the SHb% of ZB2. This result is in agreement with that of Nguyen et al. [49]. It is also reported that the addition of Mg2+ to sludge had no effect on the bound water content [57].
The effect of Al3+ on the SHb of ZB2 was significant, indicating that the estimated main effect of Al3+ on SHb was positive. Thus, AAg of ZB2 increased with increasing Al3+ (Fig. 10). Park et al. [57] reported that the concentrations of soluble and colloidal biopolymer are low when Al3+ are present in activated sludge, but improves the sludge dewatering property. This indicates that Al3+ could reduce the bound water on the surface of aggregate cells. These results suggest that Al3+ could improve the ZB2 SHb properties by increasing the cell aggregate bond.
3.2.4. The effect of cations on AAg for ZL, ZB1 and ZB2
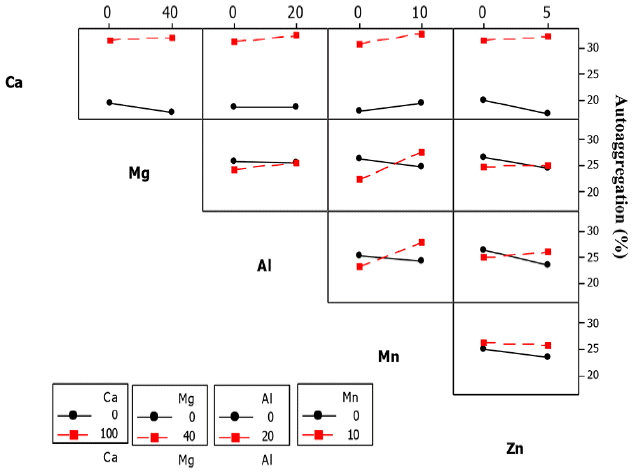
Significant 2-way interactions were observed between Al3+ and Mn2+ (Al × Mn), Al3+ and Zn2+ (Al × Zn), Mg2+ and Mn2+ (Mg × Mn) and Ca2+ and Zn2+ (Ca × Zn). The interaction between Al3+ and Mn2+ was monitored at zero Mn2+ concentration. The AAg% slightly decreased from 25% to 22% with increasing Al3+ concentration from 0–20 mg/L. The effect of cations on AAg of ZL is presented in Table 7.
Fig. 11 shows that the AAg increased in the range of 24–28% at a fixed Mn2+ and Al3+ concentrations of 10 mg/L and 20 mg/L, respectively. The combined addition of Al3+ and Zn2+ to the ZL culture at concentrations of 0 mg/L and 20 mg/L caused a slight decrease of AAg from 26% o 22%, but was in the range of 25% to 26% when Zn2+ concentration was reduced to 5 mg/L. At Al3+ and Zn2+ concentrations of 20 mg/L and 5 mg/L, AAg was about 26%.
At a Mn2+ concentration of 10 mg/L, the AAg of the ZL culture slightly decreased from 26% to 24%. AAg further increased to 29% with the addition of Mg2+ upto a concentration of 40 mg/L.
The AAg of the ZL culture decreased from 20% to 17% when Zn2+ concentration of 5 mg/L was added. However, the combined addition of Ca2+ and Zn2+ at a fixed concentrations of 100 mg/L and 5 mg/L caused an increase of AAg from 30% to 34% as shown in Fig. 11.
The interaction between Mg2+, Al3+ and Zn2+ (Mg × Al × Zn) was the only significant 3-way interactions. In the 4-way interactions, the three significant groups were Ca2+, Mg2+, Al3+ and Mn2+ (Ca2+ × Mg2+ × Al3+ × Mn2+), Ca2+, Mg2+, Mn2+ and Zn2+ (Ca × Mg × Mn × Zn) and Ca2+, Al3+, Mn2+ and Zn2+ (Ca × Al × Mn × Zn). In the 5-way interactions, Ca × Mg × Al × Mn × Zn were significant. The results of the factorial analysis for the individual and combined cationic effect on the AAg of the ZL culture varied inconsistently. The 2-way interaction effects between Ca2+ and Al3+ (Ca × Al), Ca2+ and Mn2+ (Ca × Mn), Mg2+ and Al3+ (Mg × Al), Al3+ and Mn2+ (Al × Mn) and Al3+ and Zn2+ (Al × Zn) were significant as shown in Fig. 12. The AAg of ZB1 increased from 39–43% at Al3+ concentration of 20 mg/L. An interaction effect was observed when Ca2+ concentration of 100 mg/L was added to the ZB1 culture, resulting in an increase of AAg from 50–65% (Fig. 12). In contrast, the AAg of ZB1 culture decreased from 43–39% when exposed to Mn2+ concentration of 10 mg/L, but increased from 55% to 58% when Ca2+ concentration of 100 mg/L was added to the culture (Fig. 12).
The AAg of ZB1 culture also showed an increasing trend from 41% to 59% when exposed to Al3+ concentration of 20 mg/L but slightly increased from 49% to 50% when Mg2+ concentration of 40 mg/L was added to the culture (Fig. 12). However, AAg of ZB1 culture decreased from 49% to 43% when exposed to Mn2+ concentration of 10 mg/L, but increased in the range of 50–55% when Al3+ concentration of 20 mg/L was added to the culture.
The AAg of ZB1 culture remained almost constant in the range of 46–45% when exposed to Al3+ concentration of 5 mg/L, but increased in the range of 50–55% when exposed to Al3+ and Zn2+ concentrations of 20 mg/L and 5 mg/L, respectively. In the 3-way interactions, the five significant effects observed were Ca2+, Mg2+ and Al3+ (Ca × Mg × Al), Ca2+, Mg2+ and Mn2+ (Ca × Mg × Mn), Ca2+, Mg2+ and Zn2+ (Ca × Mg × Zn), Mg2+, Al3+ and Zn2+ (Mg × Al × Zn) and Al3+, Mn2+ and Zn2+ (Al × Mn × Zn). In the 4-way interactions, the only significant interaction was between Ca2+, Mg2+, Mn2+ and Zn2+ whereas the 5-way interactions were also significant.
In the ZB2 culture, the 2-way interaction effects between Ca2+ and Al3+ (Ca × Al), Mg2+ and Al3+ (Mg × Al) and Al3+ and Mn2+ (Al × Mn) were significant. The AAg of ZB2 increased from 41–55% when exposed to Al3+ concentration of 20 mg/L, but decreased from 62–50% when Ca2+ concentration of 100 mg/L was added (Fig. 13). In contrast, AAg of ZB2 culture increased from 49–50% when exposed to Al3+ and Mg2+ concentrations of 20 mg/L and 40 mg/L, respectively and further increased to 59% when Mg2+ was excluded from the culture as shown in Table 6.
For the interaction effect between Mg2+ and Al3+, AAg of ZB1 culture slightly increased from 55–56% when exposed to Mg2+ and Al3+ concentrations of 40 mg/L and 20 mg/L, respectively but was in the range of 50–60% when Mg2+ was excluded. In the ZB2 culture, AAg decreased from 52–50% when exposed to Zn2+ concentration of 5 mg/L and further decreased from 56–54% when Al3+ concentration of 20 mg/L was added to the culture.
In the 3-way interactions, the two significant interactions were between Ca2+, Mg2+ and Mn2+ (Ca × Mg × Mn2+) and Ca2+, Mg2+ and Zn2+ (Ca × Mg × Zn) (figure not shown).
3.2.5. Interaction effect of cations on SHb for ZL, ZB1 and ZB2
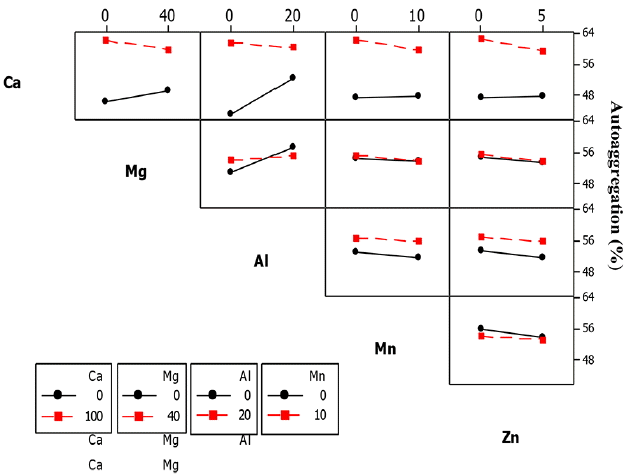
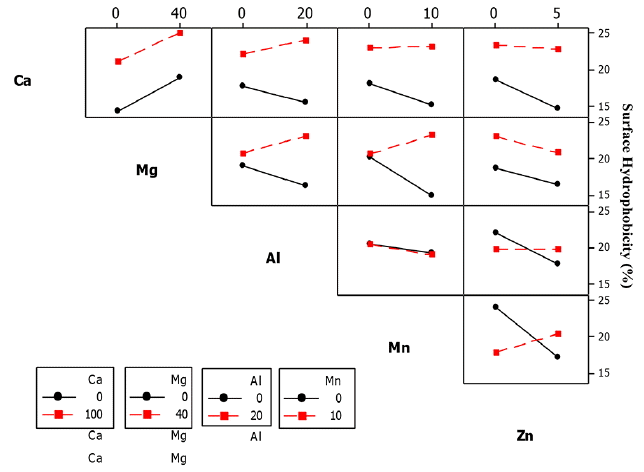
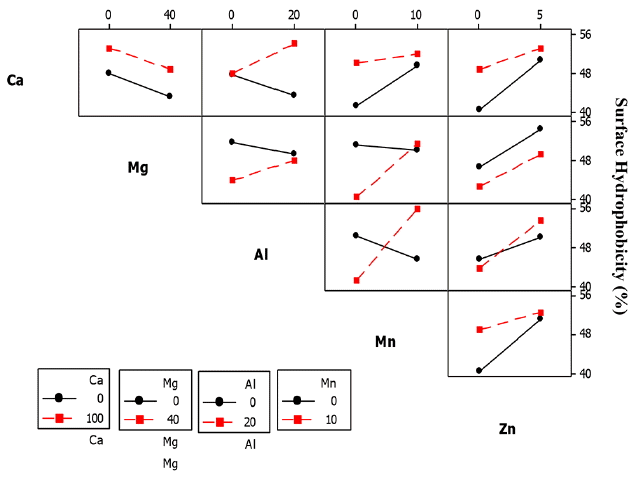
The significant 2-way interaction effect on ZL SHb was observed between Ca2+ and Al3+ (Ca × Al), Mg2+ and Al3+ (Mg × Al), Mn2+ and Zn2+ (Mn × Zn) and Mg2+ and Mn2+ (Mg × Mn) as shown in Table 8.
The SHb of ZL culture decreased from 18–15% when exposed to Al3+ concentration of 20 mg/L, but increased from 21–24% when Ca2+ concentration of 100 mg/L was added to the culture (Fig. 14). Exposure of the ZL culture to Mn2+ concentration of 10 mg/L caused a decrease of SHb from 20-14%, but showed an increasing trend from 20–24% when Mg2+ concentration of 40 mg/L was added to the culture Fig. 14.
The SHb of ZL culture also decreased from 18-14% when exposed to Zn2+ concentration of 5 mg/L and further decreased from 23-21% when Zn2+ and Mg2+ concentrations of 10 mg/L and 40 mg/L was added to the culture. The SHb further decreased from 19-14% when exposed to Al3+ concentration of 20 mg/L and further decreased from 62-50% when Ca2+ concentration of 100 mg/L was added to the culture. In the 3-way interactions of the ZL culture, the five significant interactions was between Ca2+, Mg2+ and Al3+ (Ca × Mg × Al), Ca2+, Al3+ and Mn2+ (Ca × Al × Mn), Ca2+, Al3+ and Zn2+ (Ca × Al × Zn), Mg2+, Al3+ and Zn2+ (Mg2+ × Al3+ × Zn2+), Ca2+, Mg2+ and Mn2+ (Ca × Mg × Mn), and Ca2+, Mg2+ and Zn2+ (Ca × Mg × Zn). In the 4-way interactions, two groups were significant including Ca2+, Mg2+, Al3+ and Zn2+ (Ca × Mg × Al × Zn) and Ca2+, Mg2+, Mn2+ and Zn2+ (Ca × Mg × Mn × Zn). The 5-way interactions had significant effect on SHb (figure not shown).
In the ZB1 culture, the SHb for the 2-way interactions between Ca2+ and Mg2+ (Ca × Mg), Ca2+ and Al3+ (Ca × Al), Ca2+ and Mn2+ (Ca × Mn), Ca2+ and Zn2+ (Ca × Zn) and Mn2+ and Zn2+ (Mn × Zn) were significant. The interaction effect of the cations on the SHb of ZB1 is shown in Fig. 15. The SHb decreased from 35-18% when ZB1 culture was exposed to Mg2+ concentration of 40 mg/L and further decreased from 39-37% when Ca2+ concentration of 100 mg/L was added. This showed that only Ca2+ effect influenced the SHb since addition of Mg2+ decreased the SHb. A similar decrease of SHb from 33-28% was observed when the ZB1 culture was exposed to Al3+ concentration of 20 mg/L, but increased from 37-39% when Ca2+ concentration of 100 mg/L was added (Fig. 15).
In contrast, the SHb of ZB1 culture increased from 28-32% when exposed to Mn2+ concentration of 10 mg/L, but decreased from 40-35% when Ca2+ concentration of 100 mg/L was added to the culture. The SHb slightly decreased from 36-35% when exposed to Zn2+ concentration of 5 mg/L and further decreased from 36-32% when Ca2+ concentration of 100 mg/L was added to the culture (Fig. 16). However, SHb slightly increased from 37-38% when exposed to Zn2+ and Mn2+ concentrations of 5 mg/L and 10 mg/L, respectively. In the 2-way interactions, the three significant interaction groups for SHb were Ca2+ and Al3+ (Ca × Al), Al3+ and Mn2+ (Al × Mn) and Mg2+ and Mn2+ (Mg × Mn) (Table 8). The SHb decreased from 47-42% when exposed to Al3+ concentration of 20 mg/L, but increased from 47-55% when Ca2+ concentration of 100 mg/L was added to the culture.
In the ZB2 culture, the SHb decreased from 50-45% when exposed to Mn2+ concentration of 10 mg/L, but increased from 40-56% when Al3+ concentration of 20 mg/L was added to the culture for the three way interaction (Fig. 16). SHb also decreased from 52-50% when exposed to Mn2+ concentration of 10 mg/L, but increased from 40-53% when Mg2+ concentration of 40 mg/L was added to the culture (Fig. 16).
The five significant interaction groups in the 3-way interaction effects on SHb of ZB2 were between Ca2+, Mg2+ and Al3+ (Ca × Mg × Al), Mg2+, Al3+ and Mn2+ (Mg × Al × Mn), Ca2+, Mg2+ and Mn2+ (Ca × Mg × Mn), Mg2+, Al3+ and Mn2+ (Mg × Al × Mn) and Al3+, Mn2+ and Zn2+ (Al × Mn × Zn) (Table 8). In the 4-way interactions, the two significant groups were calcium, magnesium, aluminium, and manganese (Ca2+ × Mg2+ × Al3+ × Mn2+) and calcium, magnesium, manganese and zinc (Ca2+ × Mg2+ × Mn2+ × Zn2+). The 5-way interactions also had significant effect on ZB2 SHb.
High interactions amongst the cations did not influence the AAg and SHb of microbial organisms. From the experiment and analysis, most of the cations had effects that could stimulate the microbial AAg and SHb. Literally, Ca2+ and Mg2+ were the most influential cation binder and played a vital role in microbial aggregation [23]. In biological treatment, Al3+ is the second most influential ion binder and it is known to enhance AAg of microorganisms [29, 58].
The significant effect of the combination of cations on AAg and SHb for each species of bacteria tested differed. Some of the cations such as Ca2+ had similar effects in all three bacteria. The presence of single or a mixture of cations caused different effects towards each of the species of bacteria. The result obained also showed that the presence of cations influenced the bacterial diversity for AAg and SHb. The relationship between AAg and SHb for the three bacteria was correlative. However the selection of the most performing cations could not rely on the significant effects obtained from ANOVA.
Rahman et al. [2] reported that heterogeneity of bacteria play a vital role in SHb. Heterogeneity in SHb of different bacterial strains was also reported by Del Re et al. [40]. According to Perez et al. [59], heterogeneity in SHb was due to the involvement of several factors interfering with adhesion of cells to hydrocarbons. Thus cations could be the factors that influence the heterogeneity of bacteria. Thus, different effects were observed for the various bacteria strains or species. There was good correlation between AAg and SHb in the three bacteria. This indicates that SHb and AAg results are similar and SHb could be one of the determinants of AAg. Similar to AAg, the statistical result on SHb to some extent provides an explanation with respect to the presence of cations towards SHb of cells structure and granule formation [18, 60].
4. Conclusions
The investigated input variables i.e. divalent and trivalent cations imposed significant linear effect on the selected bacterial aggregation and SHb. The ANOVA showed an overall result. Thus, Ca2+ gave the most significant effect on AAg and SHb of the selected bacteria experimented. Overall, the factorial design was purposely used in this study to test the effects of cations on the selected bacteria through the estimation of the equivalent AAg% and SHb%. In the presence of the cations especially Ca2+, ZB2 had an aggregation up to 62% which was the highest AAg amongst all of them. This was followed by ZB1 and ZL at 58% and 34%, respectively. The AAg of each bacteria correlated to the SHb.