1. Introduction
Spills are uncontrolled releases of any product, including crude oil, chemicals, or waste, into the environment. It is usually caused by equipment failure, operation mishaps, human error, or intentional damage to facilities [1]. The extent of damage depends on what, where, and how much has been spilled and how long it remains in the immediate and impacted environment [2]. An oil spill is the release of liquid petroleum hydrocarbons into the environment, especially the marine ecosystem [3]. Upon the release of oil, less volatile and heavier fractions are left behind while light fractions vaporize. Oil spill usually results in the death of aquatic and terrestrial animals and deprive the native human population of their food source and occupation [4, 5].
Cleanup of oil spill environments is laborious and may take months or even years to achieve. Different oil cleanup methods like hot water and high-pressure washing, the use of dispersants, sorbents, skimmers, oil booms, and bioremediation are currently in use [6]. Bioremediation which involves the utilization of native or introduced oil-degrading microorganisms or other forms of life to break down various components of the spilled crude in the marine environment, has become a promising innovation due to its minimal effort and eco-friendly nature [7,8]. Using this method, oil spills can be cleaned up safely, cheaply, and more efficiently than the other physical or chemical methods. Bioremediation functions basically on biodegradation, which involves the complete mineralization of organic contaminants into carbon dioxide, water, inorganic compounds, and cell protein [9].
Numerous investigations have uncovered an enormous number of hydrocarbon-degrading microorganisms in oil-rich conditions such as oil spill areas and oil reservoirs whose abundance is determined by the type of petroleum hydrocarbons and the encompassing environmental factors [10–15]. Among these organisms are Pseudomonas fluorescens, P. aeruginosa, Bacillus subtilis, Bacillus sp., Alcaligenes sp., Acinetobacter lwoffi, Flavobacterium sp., Micrococcus roseus, and Corynebacterium sp. [16]. The genus Bacillus has been reported to be an outstanding hydrocarbon degrader, and their ability to form spores when nutrients are limited makes them self-sustainable bioremediation organisms [17]. The degradation pathways of various petroleum hydrocarbons have been shown to employ oxidizing reactions; however, these pathways differ significantly because of the specific oxygenases found in different bacterial species. For instance, some bacteria can metabolize specific alkanes, while others like Proteus vulgaris and Proteus cibarius break down aromatic or resin fractions of hydrocarbons [18].
Computer-aided learning uses computer hardware, software, and networking tools to study specific subjects. The application of computational methods in the bioremediation of crude oil-polluted environments would give first-hand information about the organisms best suited for the remediation of a given crude oil pollution site since microbial enzymes degrade crude oil hydrocarbons selectively. An initial determination of the right microbes for the degradation of crude oil of a given hydrocarbon composition using in silico methods would reduce the time and cost involved in direct trials of these microbes on polluted sites. A computer-guided degradation susceptibility study of crude oil compounds on the enzyme of Bacillus subtilis protein target was studied in this paper. The findings would validate the reports on the efficiency of this organism in crude oil degradation and identify hydrocarbons most susceptible to degradation by its enzyme.
2. Materials and Methods
2.1. Collection of Crude Oil Sample
The crude oil samples were collected from Agip Oil Company Ltd, Clough Creek well No.5 ST located in the OML 63 swamp area in Bayelsa State, Niger Delta region of Nigeria. Crude oil samples were collected from the wellheads with the assistance of the field personnel of the company. Amber glass bottles with Teflon-lined lids were used for the sample collection. The collected samples were sealed and labeled and then stored in a dark insulated cooler to ensure that evaporative changes and microbial degradation did not occur in the crude oil while being taken to the laboratory. Gas chromatography analysis of the samples was done on the same day of collection.
2.2. Analysis of Crude Oil Sample
The compounds in the crude oil sample were determined using Gas Chromatography-Mass Spectrometry (GCMS-QP2010 PLUS, Shimadzu, Japan) with internal standard solution containing from n-C3 to n-C44 (prepared from aliquots of pure compounds and diluted with dichloromethane to a final concentration of 0.5 mg/mL). The chromatogram of the standard calibration solutions is presented in Fig. S1. The instrument was validated by determining the calibration curve, the limit of detection (LOD) and quantification (LOQ). By diluting certified solutions containing n-C3 to n-C44, five concentrations of calibration solutions ranging from 0.02 mg/mL to 1 mg/mL are created and used to generate the calibration curve (Fig. S1). The LOD and LOQ were determined by performing a quadruplicate blank analysis. The LOD measures the smallest quantity of the analyte, and it shows when the signal is greater than three times the noise and could be expressed as
and
meaning that the signal is ten times greater than the noise and it is the smallest amount that can be quantified. The standard deviation of the blank analysis is given as σ, and S is slope of calibration curve. The LOD and LOQ were 1.42 and 4.73 mg/mL, respectively. Following calibration, the crude oil samples were analyzed. The sample syringe was rinsed four times with the pre-solvent, four times with post-solvent, and three times with the sample. The operation conditions of the GC were as follows: carrier gas, helium (1.4 mL/min), and injector and detector temperature, 300 °C. The temperature program was 0 min at 90 °C, which was ramped to 270 °C at 6 °C/min, and held at this temperature for 30 min. The injected quantity was 1 μL of 2 % volume solution in tetrachloromethane [19].
2.3. Identification and Preparation of Ligands
The 3D structure-data files (SDF) of the compounds in the crude oil sample were identified and downloaded from the PubChem database. They were minimized in PyRx virtual screening tool, using Universal Force Field at 200 steps. They were then converted to AutoDock ligands (pdbqt) and used for the docking analysis.
2.4. Receptor Preparation
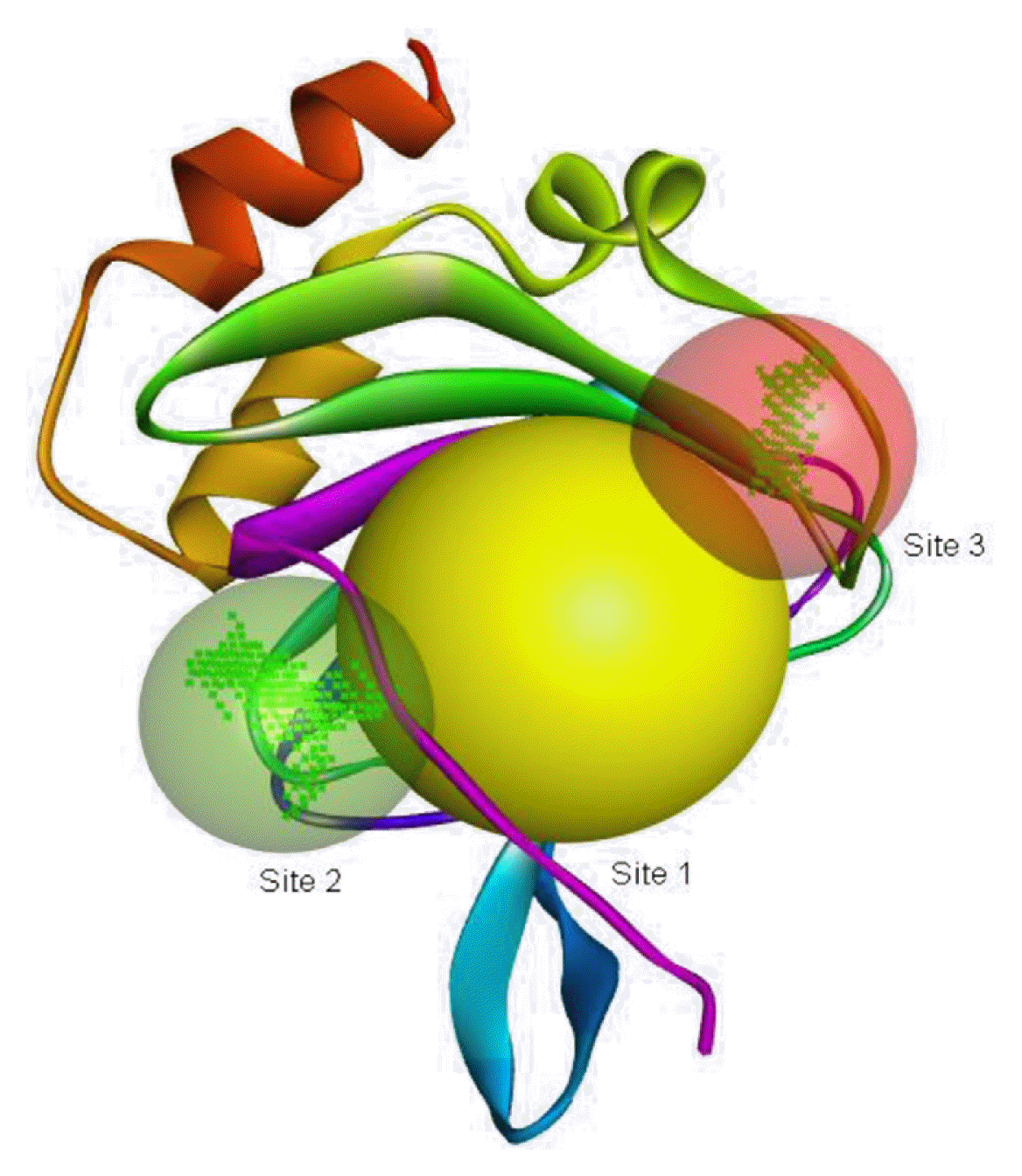
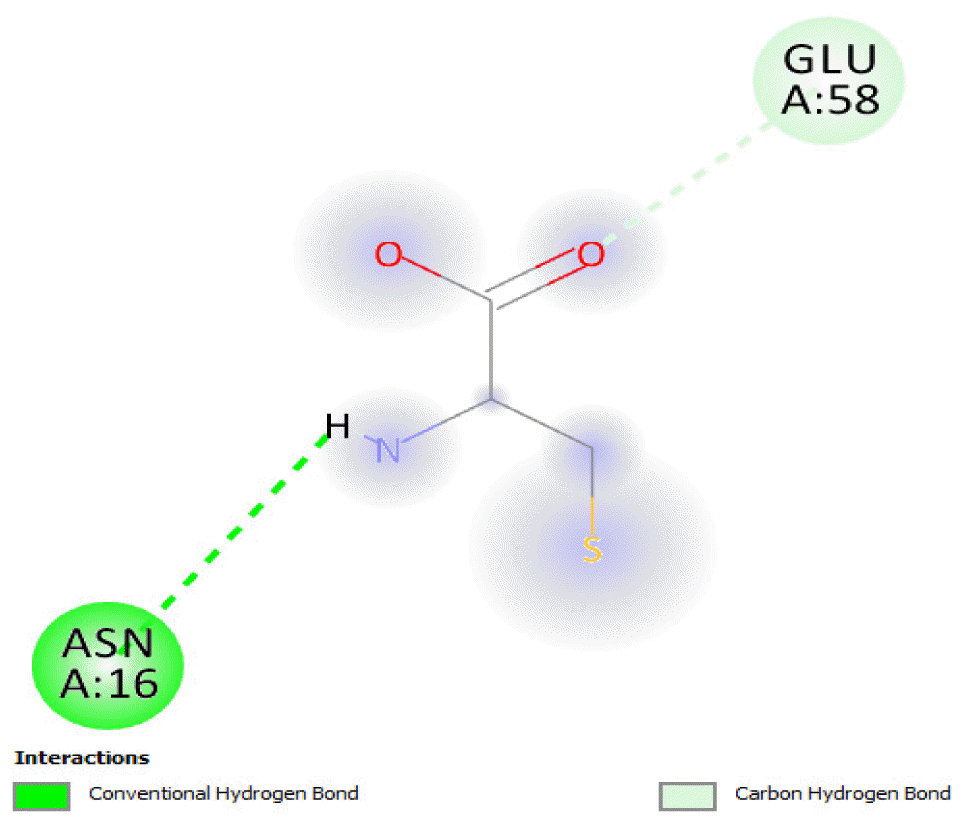
The protein of Bacillus subtilis, a putative cysteine dioxygenase (protein data band [PDB] ID: 4QM9) with resolution 2.30 Å was identified from literature [20] and used as a target in this study. Chain A of the protein was used for the docking analysis to improve the ligand-binding accuracy [21]. The interfering crystallographic water molecules and co-crystallized ligand were removed, and minimization of the energy of the protein was then done using UCSF Chimera 1.14 [22, 23]. The protein was minimized at 300 steepest descent steps at 0.02 Å. The conjugate gradient steps were 10 at 0.02 Å and 10 update intervals. Gasteiger charges were added using Dock Prep to get a good structure conformation. The active sites on the minimized protein (Fig. 1) were identified using Biovia Discovery studio 4.5, developed and distributed by Dassault Systemes BIOVIA [24].
2.5. Docking Studies
The multiple ligands docking of the crude oil compounds on the protein target was done with Autodock Vina in PyRx software version 0.8 [25,26]. Blind docking of the compounds at the protein cavities was performed to give the ligands unhindered access to interact with sites where they had minimum energy. The center grid box was set to the dimension center x: − 40.037, center y: − 18.620, center z: 142.089, and size x: 50.411, size y: 44.123, size z: 42.859. The results in terms of binding free energy (ΔG) for each compound were obtained.
3. Results and Discussion
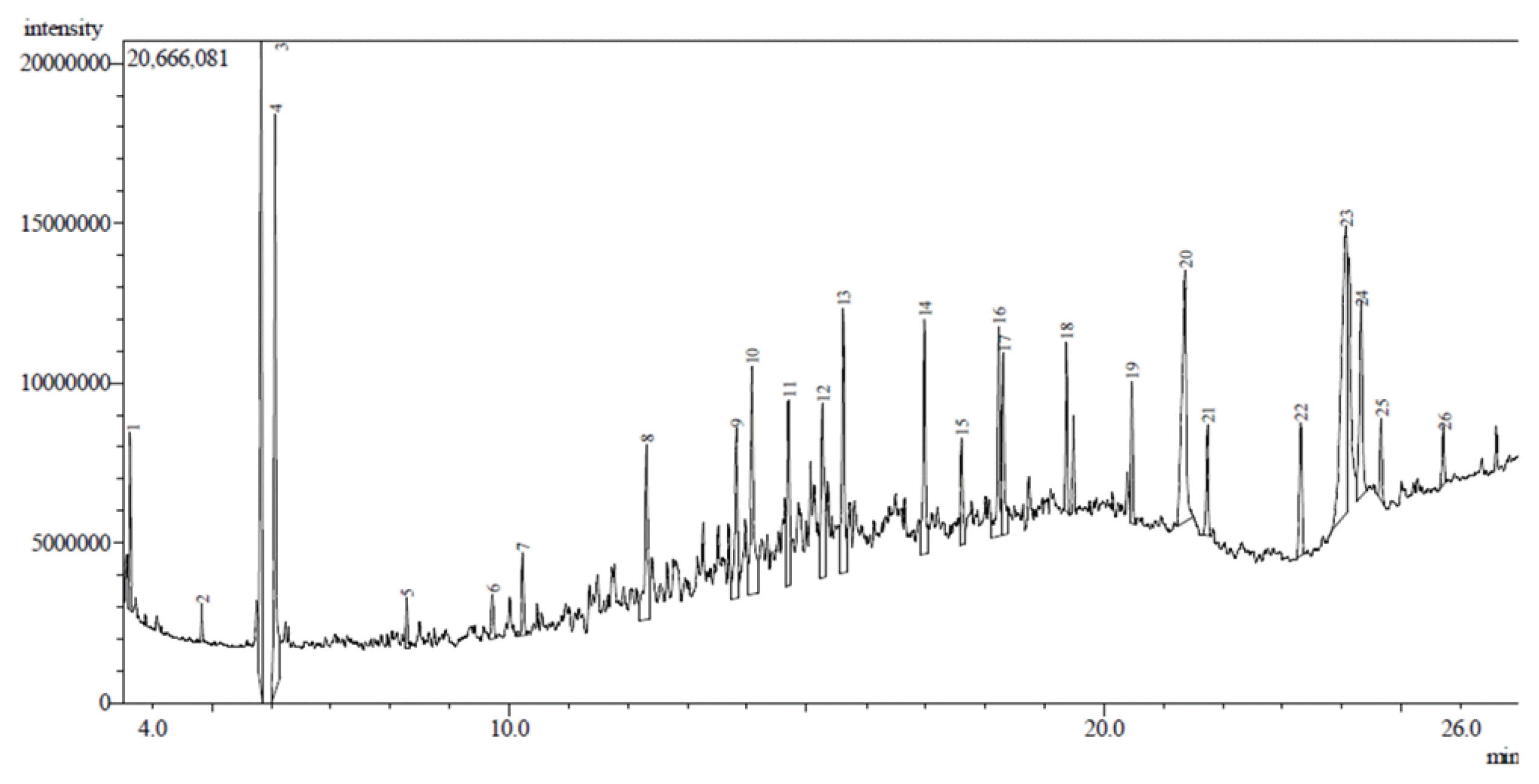
The chemical compounds eluted in the gas-chromatography analysis of the crude oil sample are shown in Fig. 2. The identified compounds and their percentage composition in the crude oil sample are summarized in Table 1. The sample was composed of 47.48 % hydrocarbons and 52.52 % hydrocarbon derivatives. The hydrocarbons comprised 29.44 % straight-chain, 13.79 % branch-chain, and 4.25 % cyclic compounds.
The hydrocarbon derivatives had 22.83 % ketones, 1.72 % alcohol, and 27.97 % carboxylic acids.
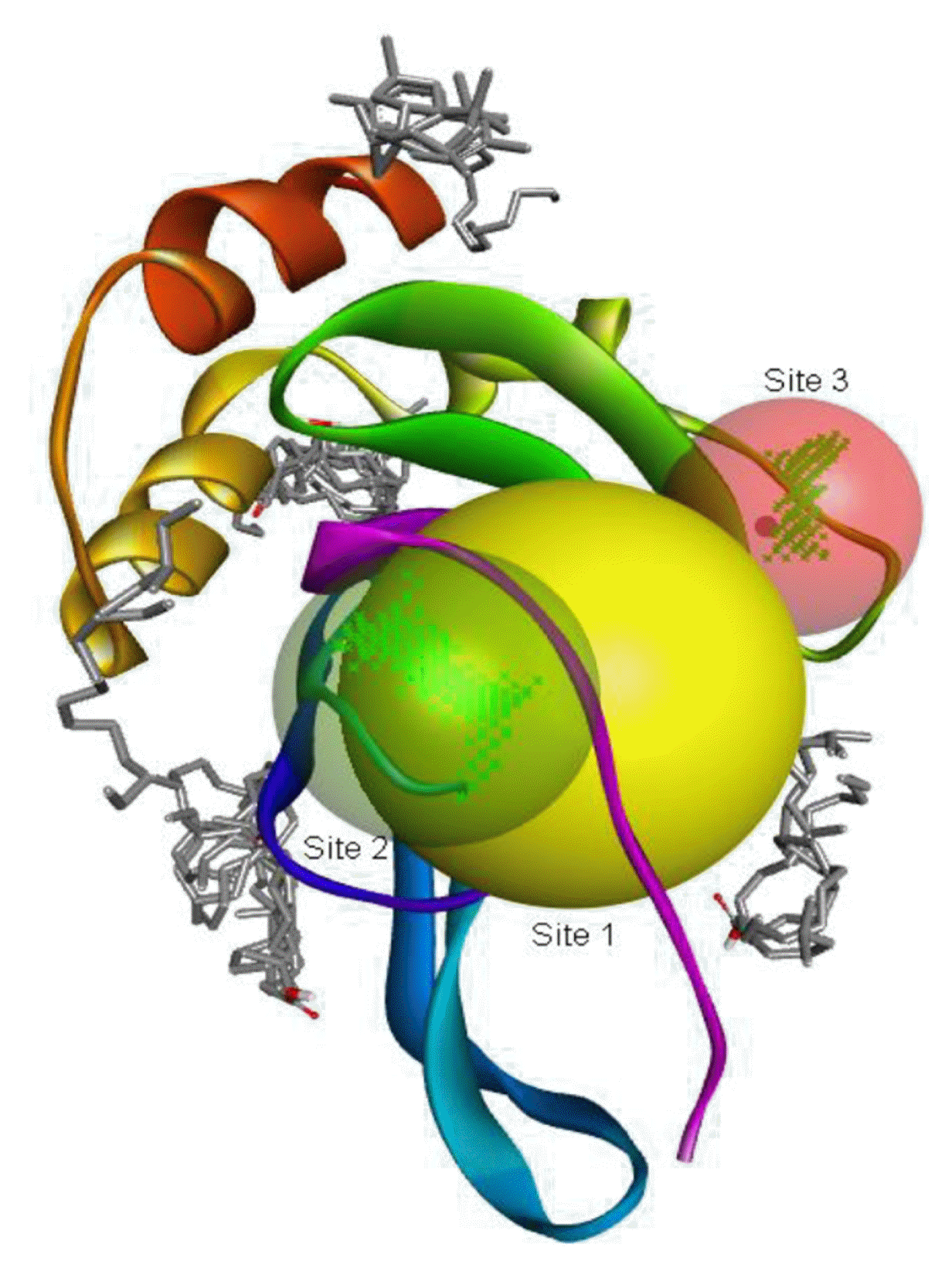
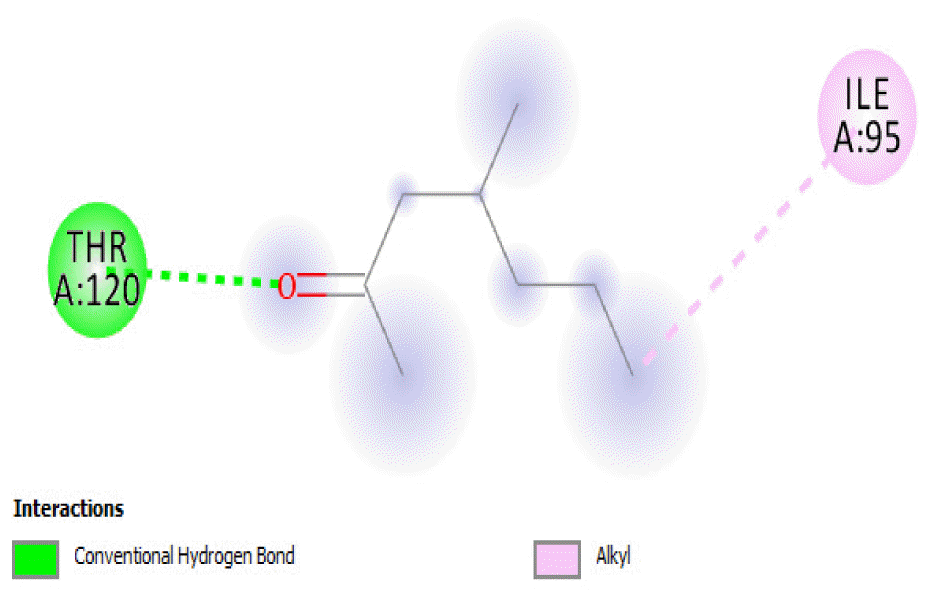
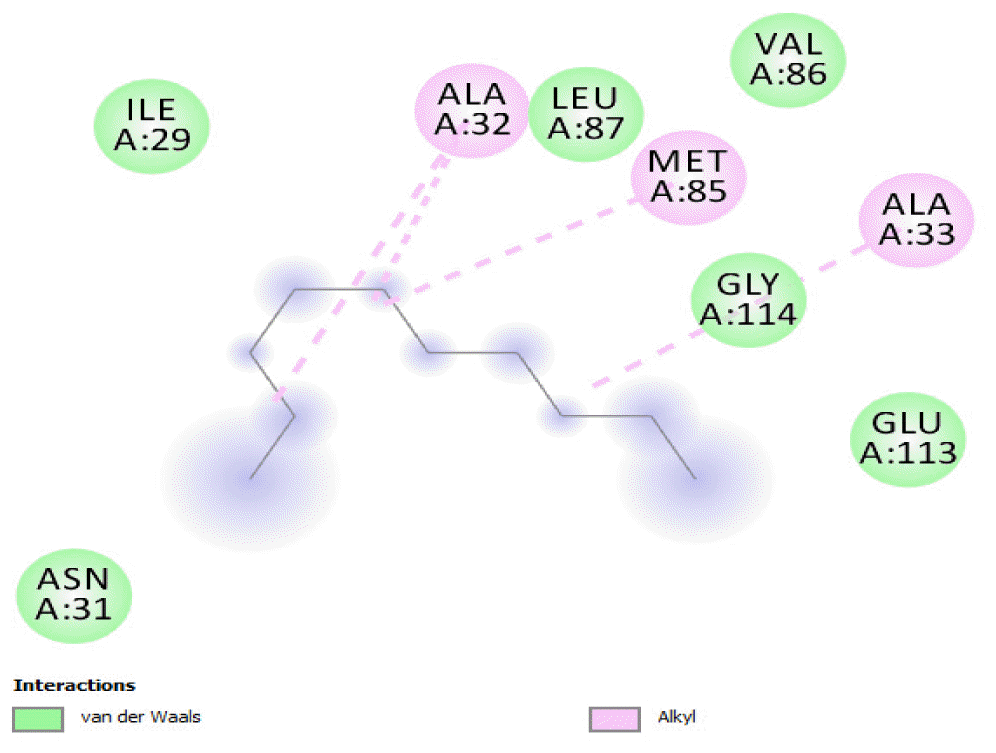
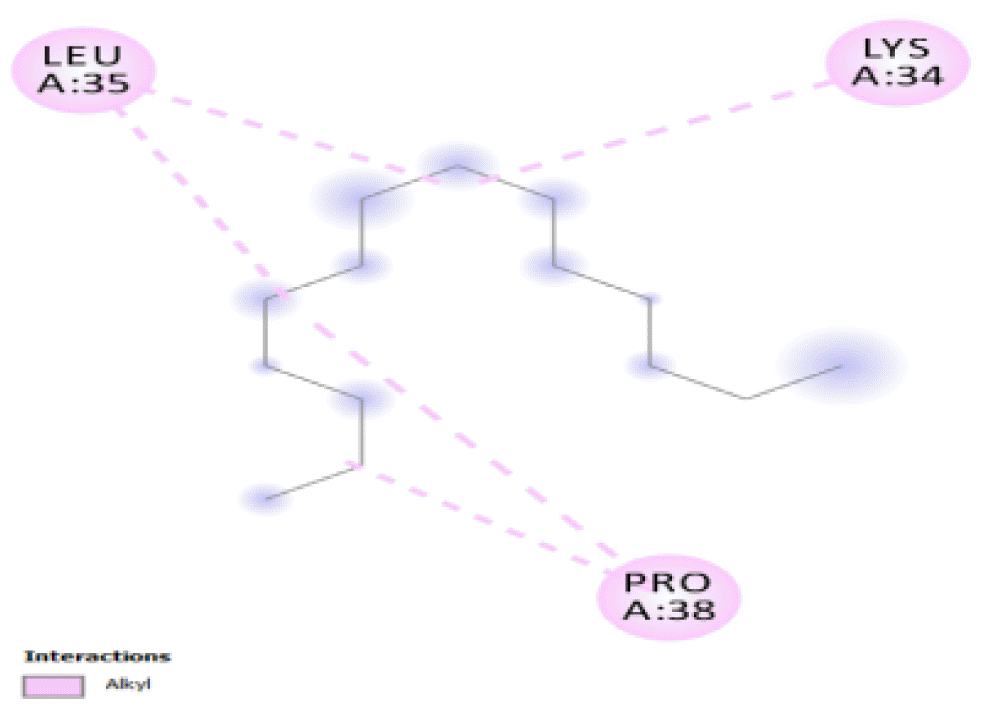
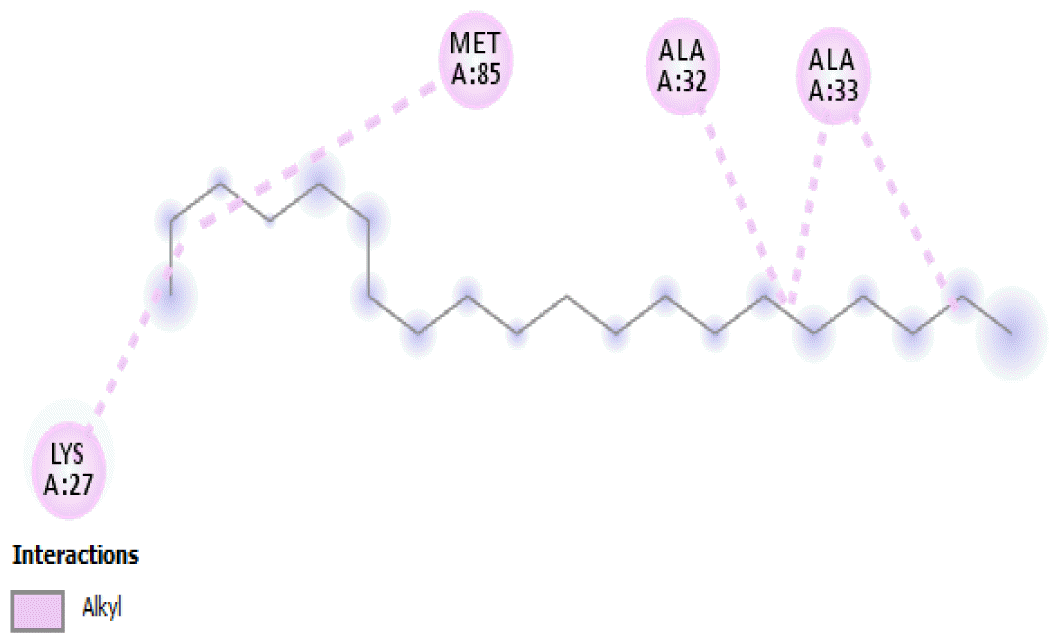
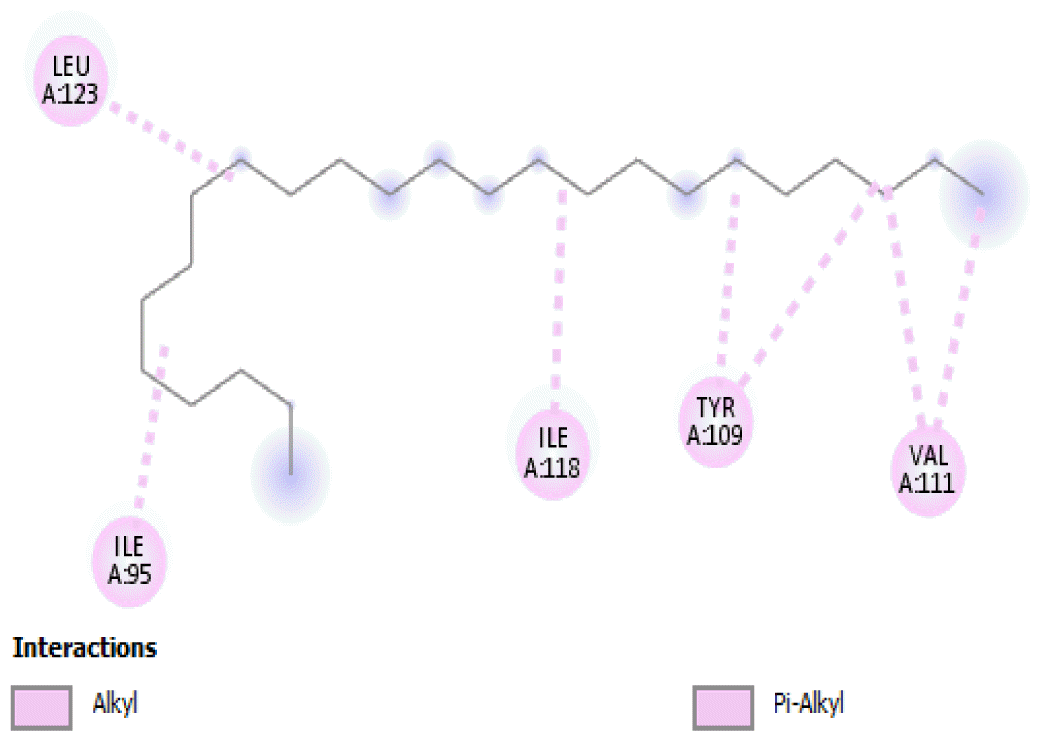
The binding positions of the hydrocarbon compounds on the protein target are shown in Fig. 3.
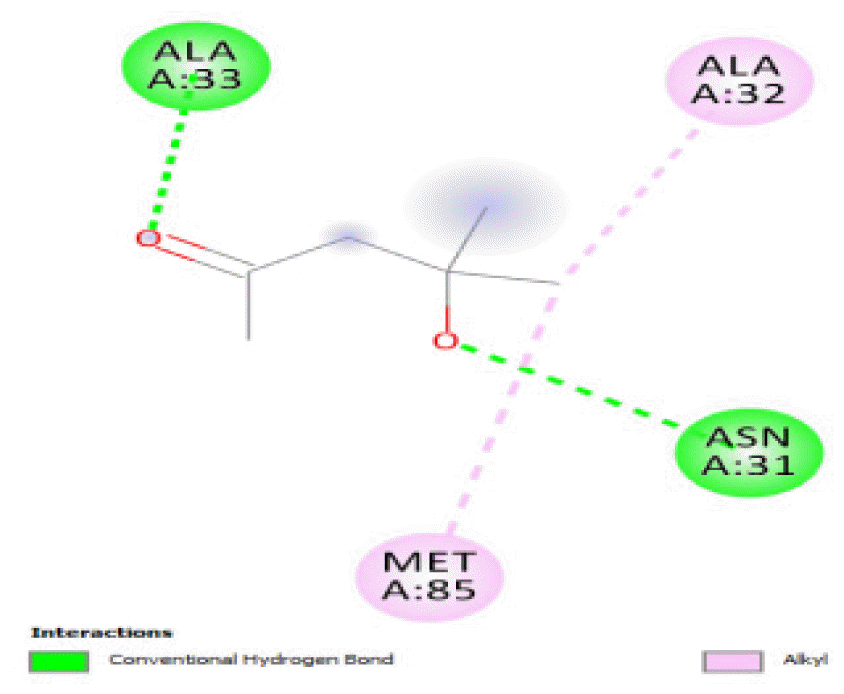
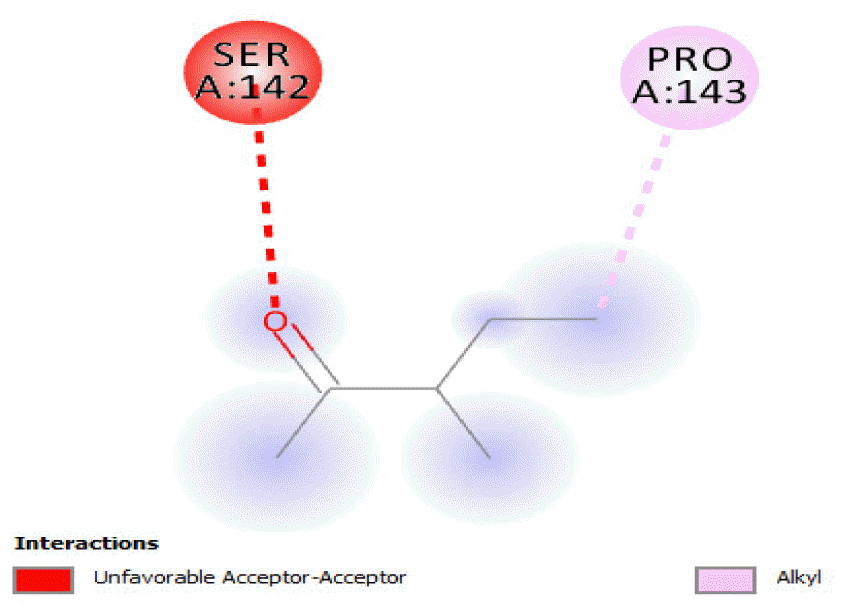
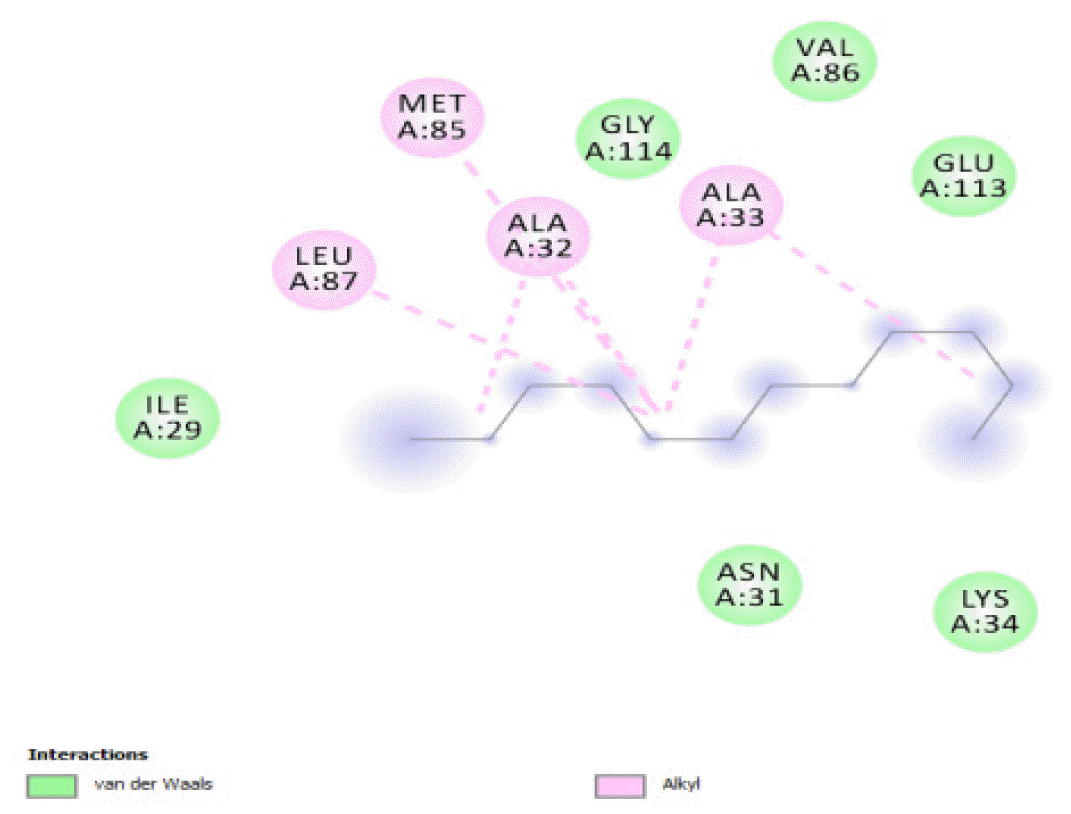
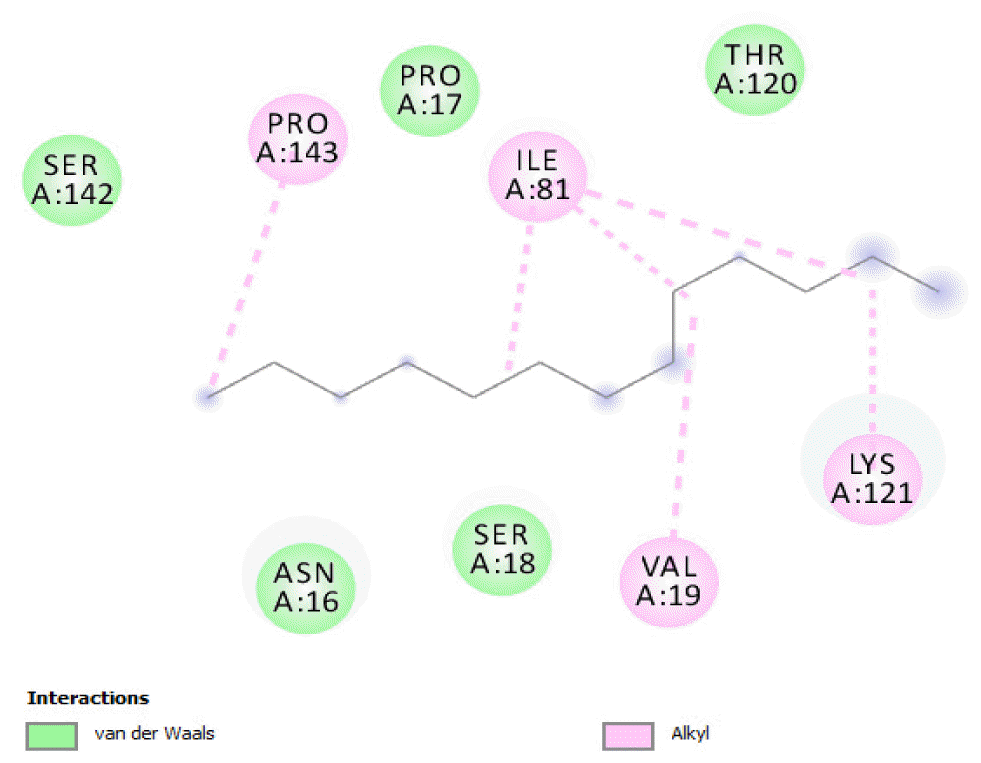
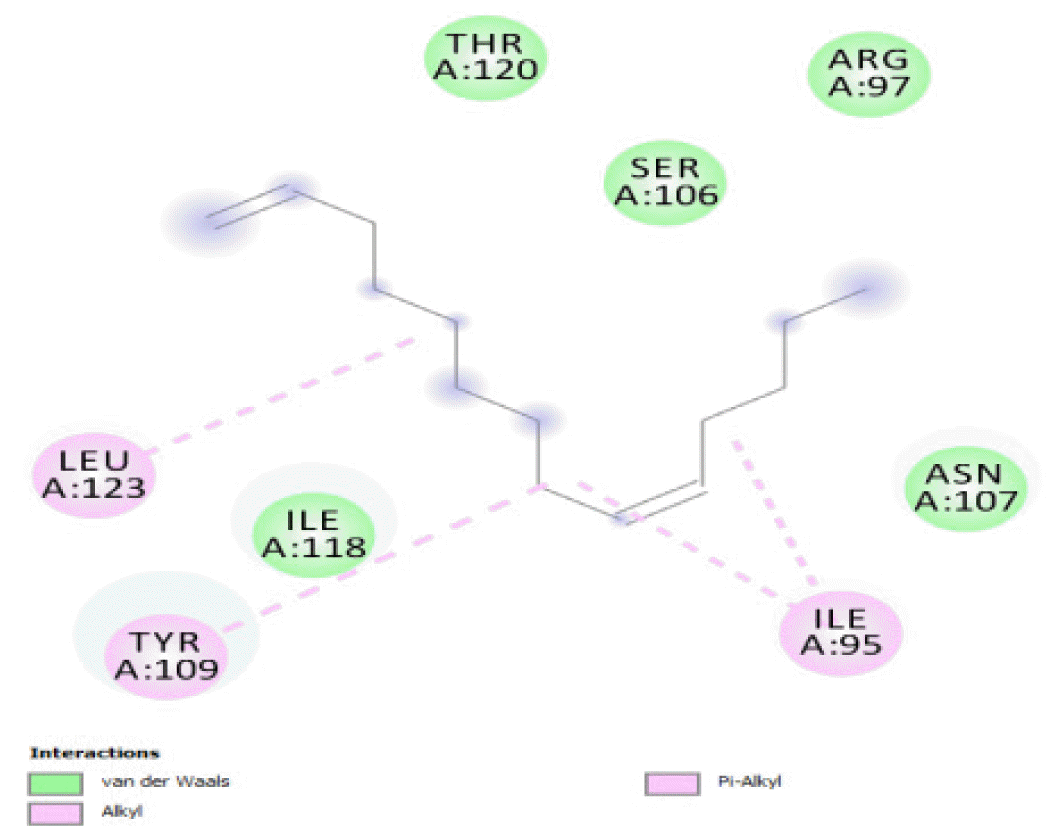
The binding of all the compounds occurred outside the active sites of the protein. Most of the compounds interacted at some specific positions on the protein target. The values of the binding free energies of the compounds on the protein are summarized in Table 2. The protein-ligand interactions of the docked hydrocarbon compounds are shown in Table 3. Hydrogen, alkyl, van der waals, pi-alkyl, and pi-sigma were the most common modes of interactions between the compounds and the protein. Most of the oxygen containing hydrocarbons as well as the control interacted with the proteins by hydrogen bonding. Alkyl interactions were found between the protein and the linear hydrocarbons, most of which had higher binding free energies, while pi-alkyl interactions were prevalent among compounds with lower free energies and the protein.
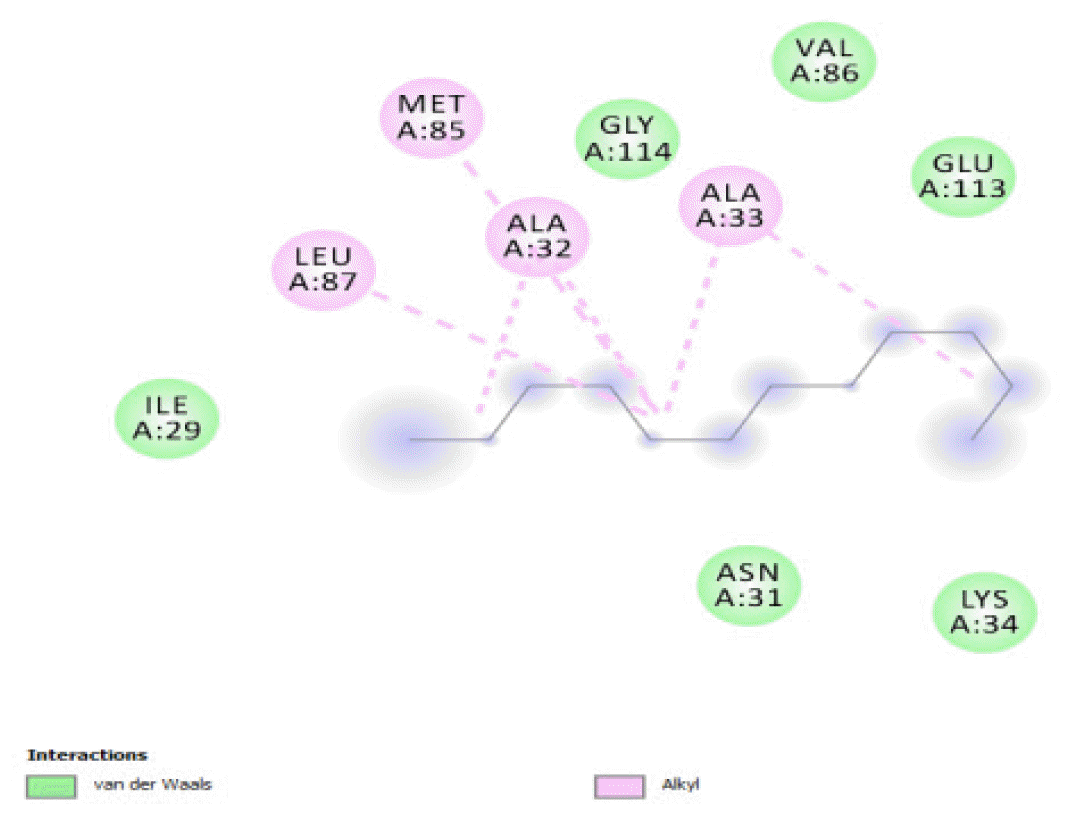
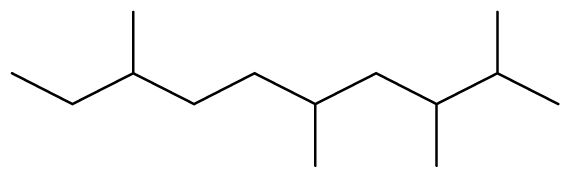
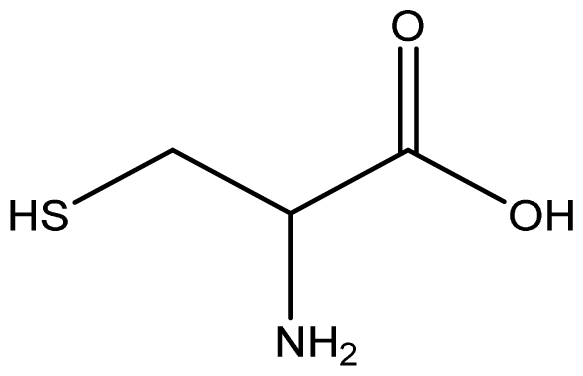
The binding free energy gives the sum of all the intermolecular interactions present between a ligand and the target. The docking score is the scoring function that predicts the binding affinity of the ligand and target after docking. The binding free energy of the co-crystallized ligand cysteine (−3.6 kcal/mol) was used as a reference standard for comparing the susceptibility of the hydrocarbons to oxidation by the protein [27, 28].The binding free energy of decane, dodecane, and eicosane was the same (−2.9 kcal/mol) and the highest for all the studied compounds. The high binding free energy values for these compounds is an indication that their binding to the protein is relatively poor. As such, their oxidation by the protein enzyme would not be feasible. The docking of the compounds also occurred at similar positions on the protein where all the compounds interacted with LYS27, ALA32, ALA33, and MET85.Tetradecane and nonadecane-2-methyl-had the same binding free energy value (−3.1 kcal/mol), the next highest set obtained. Their dock score showed that the protein enzyme would poorly oxidize them. Their binding occurred at a similar site, and both compounds interacted with LYS34.
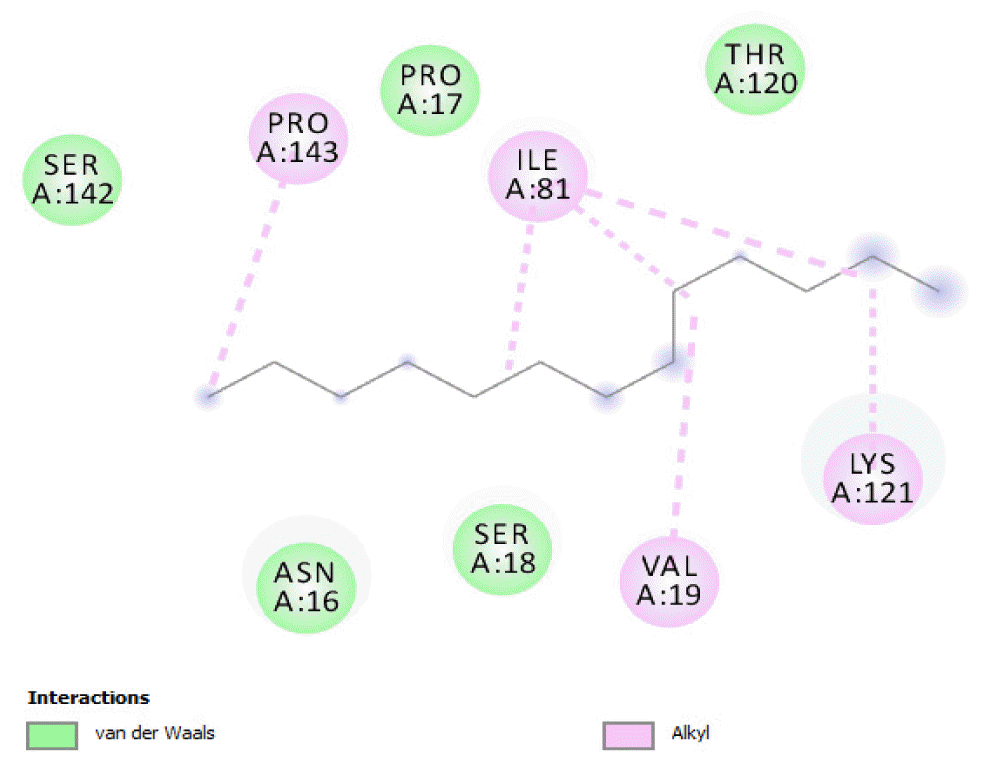
The binding free energy values of 2-pentanone 3-methyl- and 1,9-tetradecadiene were the same (−3.3 kcal/mol) and their binding occurred at different sites on the protein. The binding affinities of octane 2,4,6-trimethyl-tridecane, hexadecane, n-hexadecanoic acid, octadecane, 9-octadecenoic acid (Z)-, and n-octadecanoic acid (−3.4 kcal/mol) were also the same and their binding on the protein occurred at different sites. The oxidation of these compounds by the protein would also be relatively poor, as could be seen from their binding affinities.
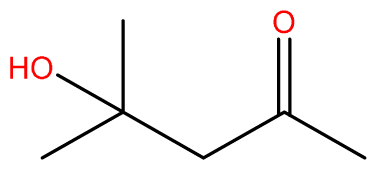
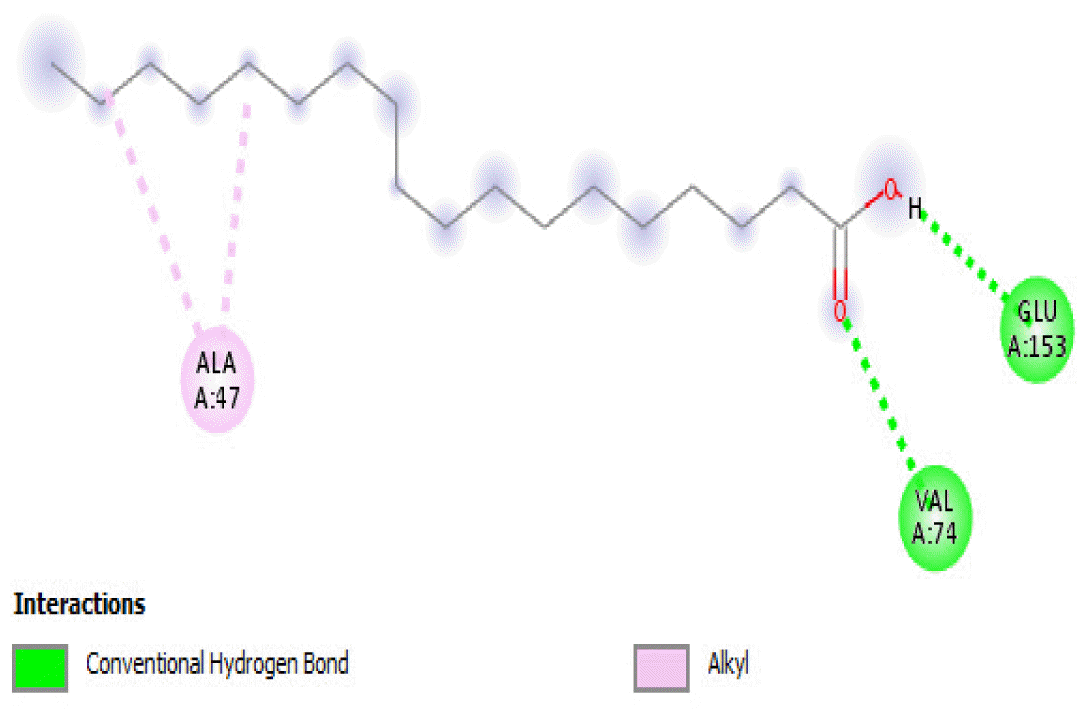
The binding free energies of 2-heptanone 4-methyl-, 4-heptanol 3-methyl-, tetracosane, and 2-pentanone 4-hydroxy-4-methyl- were close and gave −3.5 kcal/mol, −3.6 kcal/mol, −3.6 kcal/mol, and −3.7 kcal/mol, respectively. These values were the median dock scores indicating that their oxidation by the protein is very likely. The binding of 4-heptanol 3-methyl- and 2-pentanone 4-hydroxy-4-methyl-occurred at the same site, and they interacted with ALA32, ALA33, and MET85. The binding of 2-heptanone-4-methyl- and tetracosane occurred at different sites on the protein. These compounds constituted 12.52 % of the analyzed crude oil.
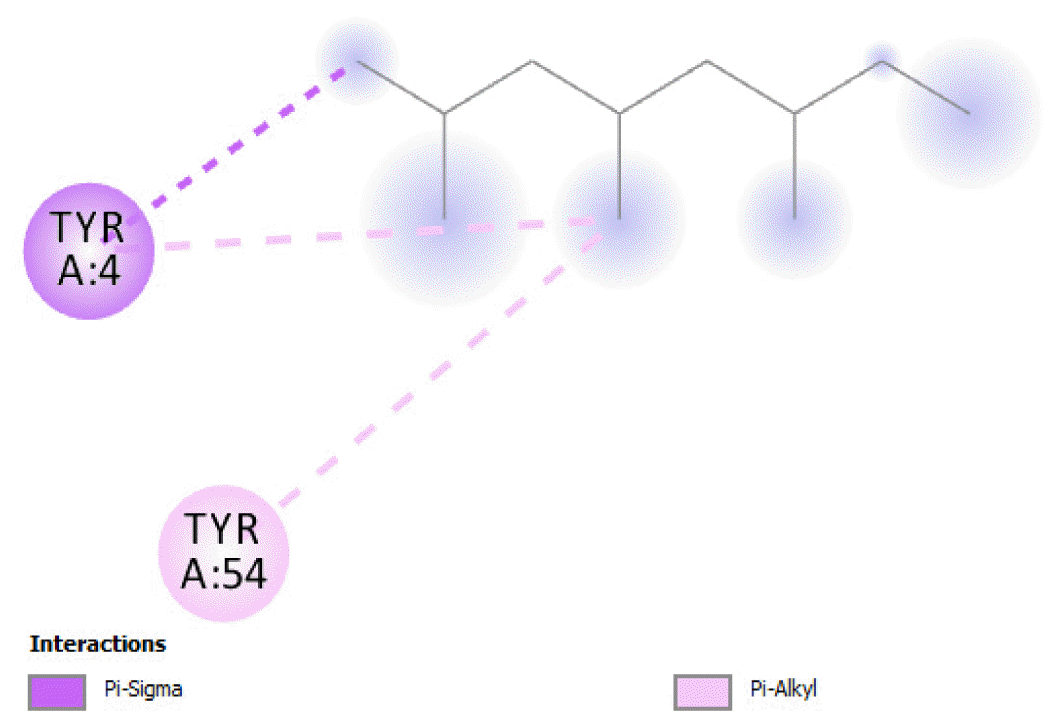
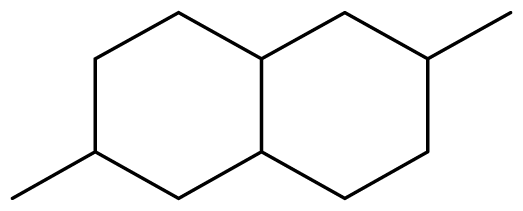
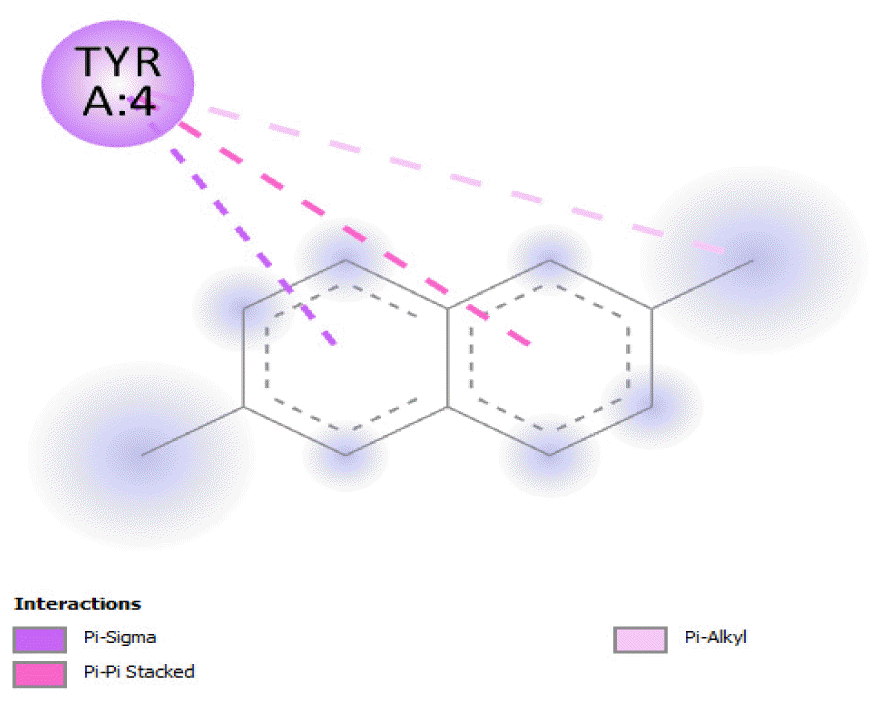
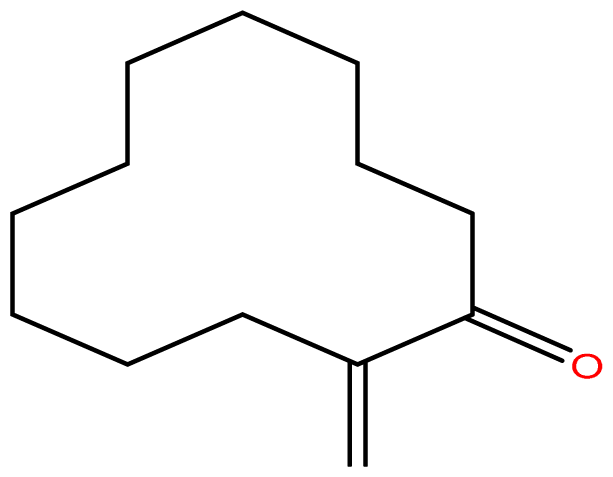
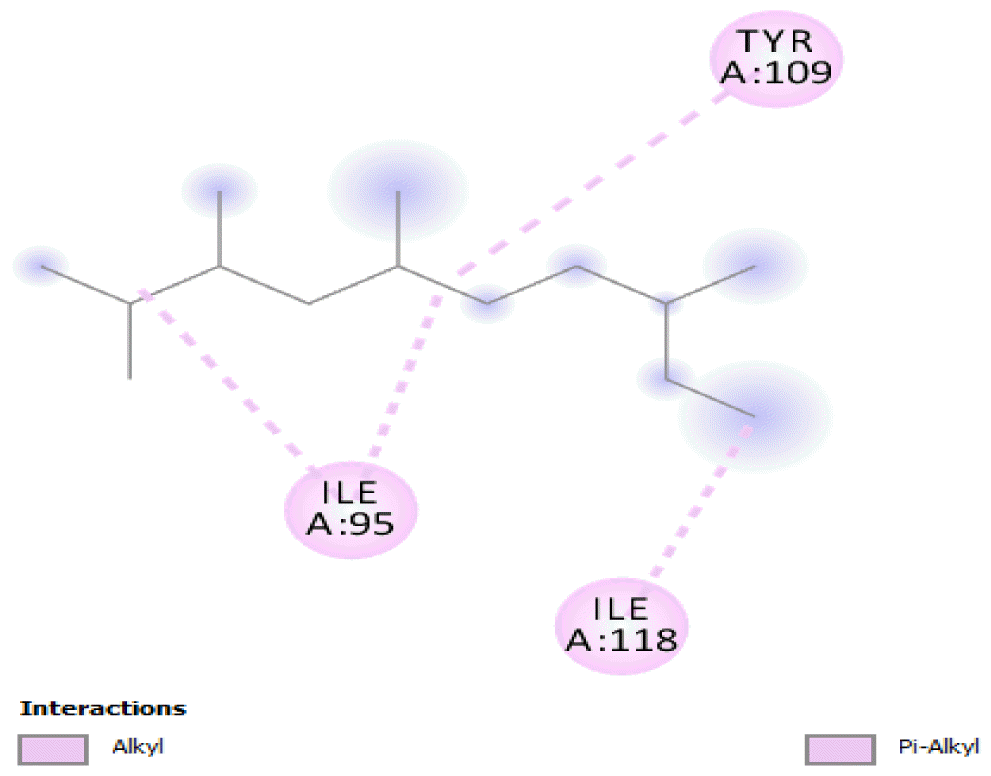
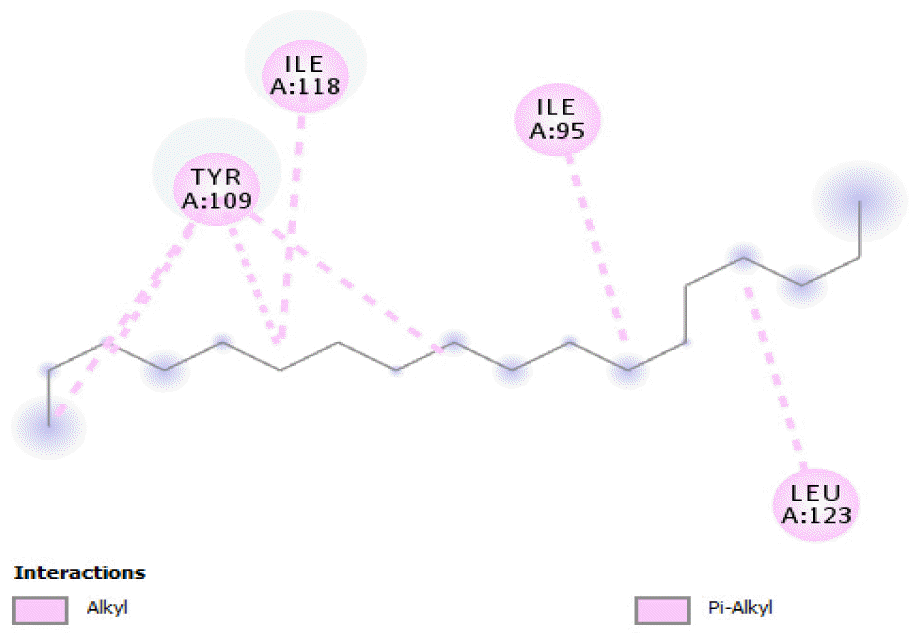
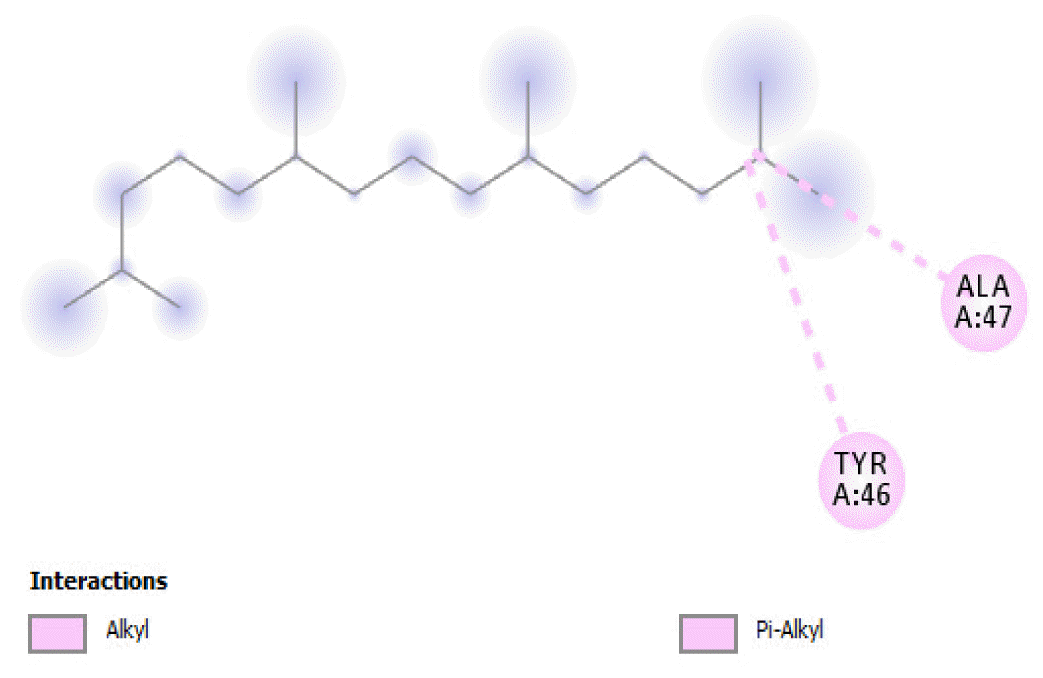
The binding free energy of naphthalene decahydro-2,6-dimethyl-, methylene cyclododecanone, decane 2,3,5,8-tetramethyl-, 1H-indene octahydro 2,2,4,4,7,7-hexamethyl-trans-, pentadecane 2,6,10-trimethyl-, and pentadecane 2,6,10,14-tetramethyl-were −4.8 kcal/mol, −4.8 kcal/mol, −3.8 kcal/mol, −5.1 kcal/mol, −3.8 kcal/mol, and −4.0 kcal/mol respectively and were relatively very good. The binding of naphthalene decahydro 2,6-dimethyl-and 1H-indene octahydro-2,2,4,4,7,7-hexamethyl-trans-occurred at the same site, and they interacted with TYR4. Methylene cyclododecanone, decane 2,3,5,8-tetramethyl-, and pentadecane 2,6,10-trimethyl-binding occurred at the same site and interacted with TYR109. Binding of pentadecane 2,6,10,14-tetramethyl-occurred in a pocket different from all the other compounds in this classification, and it interacted with the residues TYR46 and ALA47. These compounds constituted 15.39 % of the analyzed crude oil and are either cyclic or highly branched. These observations suggested that compounds of this structural nature are more susceptible to oxidation by B. subtilis and would be efficiently remediated in environments polluted by crude oil.
The data from this study affirms earlier reports that crude oil hydrocarbons have different affinities on enzyme pockets. With advancements in the genetic engineering of different microbes to enhance their crude oil remediation potentials, in silico techniques would become a veritable tool for costless validation studies of the efficiency of the modified microbial enzymes in degrading specific hydrocarbons. Genetically engineered microbes are used in bioremediation to design novel strains that can degrade high molecular weight polyaromatic compounds [29]. Data from molecular docking of hydrocarbons on enzyme surfaces could be used to identify particular sites on the enzyme where degradation occurs, the mechanism of the degradation process, and possible positions on binding sites where modifications are required to increase the binding affinity of specific crude oil hydrocarbons at those sites before their actual implementation.
4. Conclusions
The selectivity of hydrocarbons in crude oil by the cysteine dioxygenase enzyme of Bacillus subtilis was studied in silico. The crude oil sample used for the study contained 47.48 % hydrocarbons and 52.52 % hydrocarbon derivatives. The binding free energy values of the compounds on the protein target indicated that most of the alkanes would not be easily oxidized by the bacterial enzyme as shown by their high binding energy (−2.9 kcal/mol), while poly-branched and cyclic hydrocarbons with binding energies in the range −3.8 kcal/mol to −5.1 kcal/mol would undergo this process more rapidly. These findings indicate that B. subtilis would not achieve complete oxidation of all hydrocarbons and hydrocarbon derivatives in crude oil polluted environments independently.