The potential of common duckweed (Lemna minor) in phytoremediation of phenanthrene and pyrene
Article information

Abstract
The distribution of polycyclic aromatic hydrocarbons (PAHs) as a group of toxic and persistent aromatic pollutants in the environment is rapidly enhancing. These compounds have adverse impacts on the health of living organisms. Hence, in the present study, we investigated the potential of duckweed (Lemna minor) as an aquatic plant species for uptake, accumulation, and biodegradation of phenanthrene and pyrene under controlled conditions. L. minor plants were treated with 10 and 20 mg L−1 concentrations of phenanthrene and pyrene at the experimental duration of ten days. According to the results obtained, the toxicity of phenanthrene and pyrene contaminants on L. minor was influenced by the different initial PAHs concentrations. An increase in phenanthrene and pyrene concentration significantly decreased all studied growth parameters such as fresh weight, dry weight, and RFN and also photosynthetic pigment contents of the plant. Phenanthrene and pyrene concentrations were measured using high-performance liquid chromatography (HPLC) technique after 10 days of exposure to the PAHs. The results revealed that L. minor species could bioaccumulate effectively both typical PAHs. Furthermore, the gas chromatography-mass spectroscopy (GC/MS) technique explained the biological degradation of phenanthrene and pyrene by L. minor in the present research, and accordingly, several intermediate by-products were identified.
1. Introduction
During recent decades, polycyclic aromatic hydrocarbons (PAHs) have become one of the largest environmental pollutants groups in many studies [1–4]. PAHs are composed of at least two fused benzene rings [5]. These compounds are released into the environment, such as soil, air, sediments, and water, either by natural [6] or anthropogenic activities [7]. Even in a low concentration, PAHs have harmful effects on the environment, living organisms, and human health due to their toxic, mutagenic, and carcinogenic properties [8, 9]. Since PAHs contaminated many sites, ecologists have found remediation challenging [10].
Different physicochemical and biological techniques have been developed to preserve the ecological stability of the ecosystems against PAHs [11]. Recently, phytoremediation, which deals with applying plants for environmental remediation, has emerged as one of the promising efficient, low-cost, eco-friendly biological approaches [12–14]. The green plants can be introduced as a pollution cleanup tool in a terrestrial and aquatic environment, gaining a surge of interest in recent decades [15, 16]. Over the years, extensive literature demonstrated the considerable potential of plants in removing various pollutants in ecosystems, including aquatic ones [17–22]. In this case, some studies showed that polycyclic aromatic hydrocarbons, including pyrene and phenanthrene, are toxic compounds for almost all organisms. However, some plants can uptake them by roots and translocate them to various aerial parts. For example, maize plants absorbed and translocated these compounds to the roots and shoots with different ratios and metabolized them. Although very high concentrations have shown toxic effects on the plants [1]. Among several plant species, L. minor has been proposed as an appropriate biosystem for phytoremediation of pollutants in contaminated water resources due to its specific vital features, including high multiplication rates, widespread and small genome size, and vegetative propagation [12, 23, 24]. L. minor has been applied separately or in combination with other aquatic plants as an efficient bioaccumulator of various chemical pollutants in surface waters [23, 25]. However, scientists paid little attention to the potential of L. minor in the biodegradation of organic pollutants such as PAHs.
In the present study, we examined the potential of L. minor for phytoremediation of two typical PAHs, phenanthrene and pyrene, as the representative of 3-ring and 4-ring PAHs, respectively [3, 26]. US Environmental Protection Agency (US-EPA) put phenanthrene and pyrene in the priority pollutants list for monitoring the environment [7, 27]. The changes in some biochemical parameters, including photosynthetic pigment content and growth, were used to determine the toxic effects of phenanthrene and pyrene on duckweed plants. Subsequently, their entrance and concentrations into L. minor were investigated using the HPLC technique. Moreover, the biological degradation by-products formed in the present process were identified and characterized by the GC-MS method.
2. Material and Methods
2.1. Plant Collection and Culture Conditions
L. minor plants were collected from rice fields in Sari County, north of Iran. The collected plants were washed with distilled water to remove unwanted organisms, sterilized in the laboratory by 0.5% sodium hypochlorite for 2 min, and transferred into the glass aquarium containing 50% Steinberg culture medium. The substances of this culture medium were purchased from Merck Co. (Germany) and contained 349 mg L−1 KNO3 (≥ 99%), 12.5 mg L−1 K2HPO4 (≥ 98%), 90 mg L−1 KH2PO4 (≥ 99.5%), 108 mg L−1 Ca(NO3)2.4H2O (≥ 98%), 0.04 mg L−1 NaMoO4.2H2O (99.5%), 49 mg L−1 MgSO4.7H2O (98%), 0.1 mg L−1 MnCl2.2H2O (99%), 0.456 mg L−1 FeCl3.7H2O (99%) 0.1 mg L−1 ZnSO4.7H2O (≥ 99%), 1.1 mg L−1 H3BO3 (99.5%) and 1.5 mg L−1 Na2EDTA(≥ 99%) [18]. Photoperiod and room temperature were regulated at 16/8h (light/dark) and 20–25°C, respectively. The culture medium containing the nutrient solution was aerated continuously with aquarium pumps and replaced every week for the optimum growth of plants.
2.2. Experimental Design
We prepared fifteen 250-mL conical flasks containing 100 mL of 50% Steinberg culture medium to set up the experimental design. After the preliminary experiments to find relevant and tolerable concentrations of phenanthrene (≥ 97%, Merck, Germany) and pyrene (≥ 96%, Merck, Germany), two engagements, including 10 and 20 mg L−1 (using a 1,000 mg L−1 stock solution of PAHs in acetone), were carefully chosen for the treatment of L. minor plants. After completely evaporating acetone (≥ 99%, Merck, Germany), L. minor plants were exposed to the culture medium for ten days and used for different assays. The experiences were repeated three times for control (free-PAHs solution containing plant) and treated samples.
2.3. Evaluation of Growth Rate
The growth rate of L. minor plants treated by 10 and 20 mg L−1 concentrations of PAHs was estimated by relative frond number (RFN), fresh weight, and dry weight. Forty fronds with relatively equal sizes and 0.1 g of the initial fresh weight were placed into each concentration for fresh and dry weight measurements. In addition, we used 20 isometric fronds with 0.5 g of the initial fresh weight for RFN measurements. After ten days, the samples’ fresh and dry weight (after drying in the oven at 50°C for 24 h) were measured. RFN of L. minor plants was determined by counting the fronds over ten days on days 0, 2, 4, 6, 8, and 10. RFN was calculated as follows [28]:
N1 and N0 represent frond numbers at day N and 0, respectively.
2.4. Evaluation of Photosynthetic Pigments
The fronds were treated with 10 and 20 mg L−1 concentrations of PAHs for ten days. Then, a 0.1 g fresh sample was homogenized in 100 % acetone to determine plant photosynthetic pigment contents [Chlorophyll a (Ca), chlorophyll b (Cb), and total carotenoids (Cx+c). The homogenates were centrifuged at 5,000 rpm for 10 min, and the absorbance of the supernatant was evaluated spectrometrically at 470, 645, and 662 nm. The amounts of photosynthetic pigments were measured as described by Lichtenthaler (1987) [29].
2.5. Phytoremediation Estimation
After 10 days, L. minor samples treated with 10 and 20 mg L−1 concentrations were harvested from culture media and homogenized in 4ml of 2-propanol (99.5%, Merck, Germany) using mortar and pestle to extract PAHs. Then 10 ml of n-hexane (95%, Merck, Germany), as a solvent, was added to the homogenates and thoroughly mixed. The n-hexane containing the organic compounds was vaporized, and the remaining material was dissolved in 1 mL of absolute methanol (99.9%, Merck, Germany) and concentrated [30]. Phenanthrene and pyrene were quantified using high-performance liquid chromatography with a fluorescence detector and a Φ 4.6 × 150 mm reverse phase C18 column. The fluorescence detector wavelength was set at 260 nm, and the methanol was used as the mobile phase with a flow rate of 1 mL min−1. The column temperature was 30°C, and the whole running time for each injection was set to 8 min.
GC-MS analysis of the organic extracts containing 20 mg L−1 of PAHs was performed to identify the intermediate products resulting from biodegradation of PAHs by L. minor. The study of samples was carried out using an Agilent 7890 A gas chromatography with a 30-m, 0.25 μm HP-5MS capillary column coupled with an Agilent 5975 D mass spectrometer operating in electron ionization (EI) mode at 70 eV and a mass/charge range of 40–550 m/z. Helium (1.0 mL min−1) was used as the carrier gas. The oven temperature was programmed to hold at 40°C for 5 min initially, ramped up at 8°C min−1 to 280°C and saved at 280°C for 10 min. The temperatures of the inlet and detector were set at 260°C and 280°C, respectively.
2.6. Statistical Analysis
The data with three replicates were subjected to analysis of variance (ANOVA) using Duncan’s multiple comparison test by SPSS version 21 software. Significance was considered statistically at p < 0.05. The measurements were expressed as the mean ± standard deviation.
3. Results and Discussion
3.1. Effect of PAHs on the Growth of the Plant
The plant’s growth was reduced significantly in response to 10 days of exposure to 10 and 20 mg L−1 of PAHs (Fig. 1). When plants were treated with 10 and 20 mg L−1 of phenanthrene, RFN was decreased by 82.5% and 49.1%, respectively (Fig. 1(a)). Also, both pyrene concentrations had negative influences on RFN (72.35% and 67.01%, respectively, compared to the control) (Fig. 1(b)). In addition, ten days after exposing the plants to PAHs, a notable decline was observed in fresh weight (Fig. 1(c), (d)) and dry weight (Fig. 1(e), (f)) by two concentration values (10 and 20 mg L−1) of PAHs in comparison to control sample (P < 0.05). Altogether, 20 mg L−1 of phenanthrene and pyrene demonstrated a more substantial inhibitory effect on the growth of L. minor. This finding confirmed previous observations, showing the toxicity of PAHs on some species of aquatic organisms as well as maize plants that had a significant decrease in shoot and root length, fresh and dry weight with 25 to 100 ppm of pyrene and phenanthrene [31–33]. This toxic inhibition should be a consequence of photosynthesis constraints or oxidative stress as a defense response of plants to continuous stressful conditions [34].

The effect of various concentrations of phenanthrene and pyrene on Lemna minor by the means of RFN, on the 2nd, 4th, 6th, 8th and 10th days after treatment (a), (b) and fresh weight (c), (d) and dry weight (e), (f)) after 10 days of exposure. Different letters indicate significant differences (P < 0.05) according to Duncan Test (mean ± SD, n = 3)
3.2. Effect of PAHs on Photosynthetic Pigment Contents
As shown in Fig. 2, after a 10-day exposure of the plants to 10 and 20 mg L−1 of phenanthrene and pyrene, the content of the chlorophyll a, chlorophyll b, total chlorophyll, and carotenoids significantly decreased. Accordingly, a considerable reduction was shown in the chlorophyll a content (26.5% and 17.5%), the chlorophyll b content (42% and 30%), and carotenoids (35% and 28%) in 20 mg L−1 phenanthrene and pyrene treatments in comparison with the control sample, respectively (Fig. 2(a), (b)). In a similar trend, adverse effects of PAHs on photosynthetic pigment contents were previously demonstrated in other plant species and algae. For example, the photosynthetic pigments content of C. vulgaris gradually decreased with the enhancing concentration of phenanthrene from 2 to 50 ppm. Also, the time and concentration-dependent impact of PAHs caused a decrease in total chlorophyll and protein content on C. vulgaris [35, 36]. The photosynthetic pigment contents reduction in plants is considered a physiological biomarker of pollutants phytotoxicity [37]. Therefore, it could be concluded that phenanthrene and pyrene declined the photosynthesis capacity of L. minor that may be a protective response to limit reactive oxygen species (ROS) production in chloroplasts.
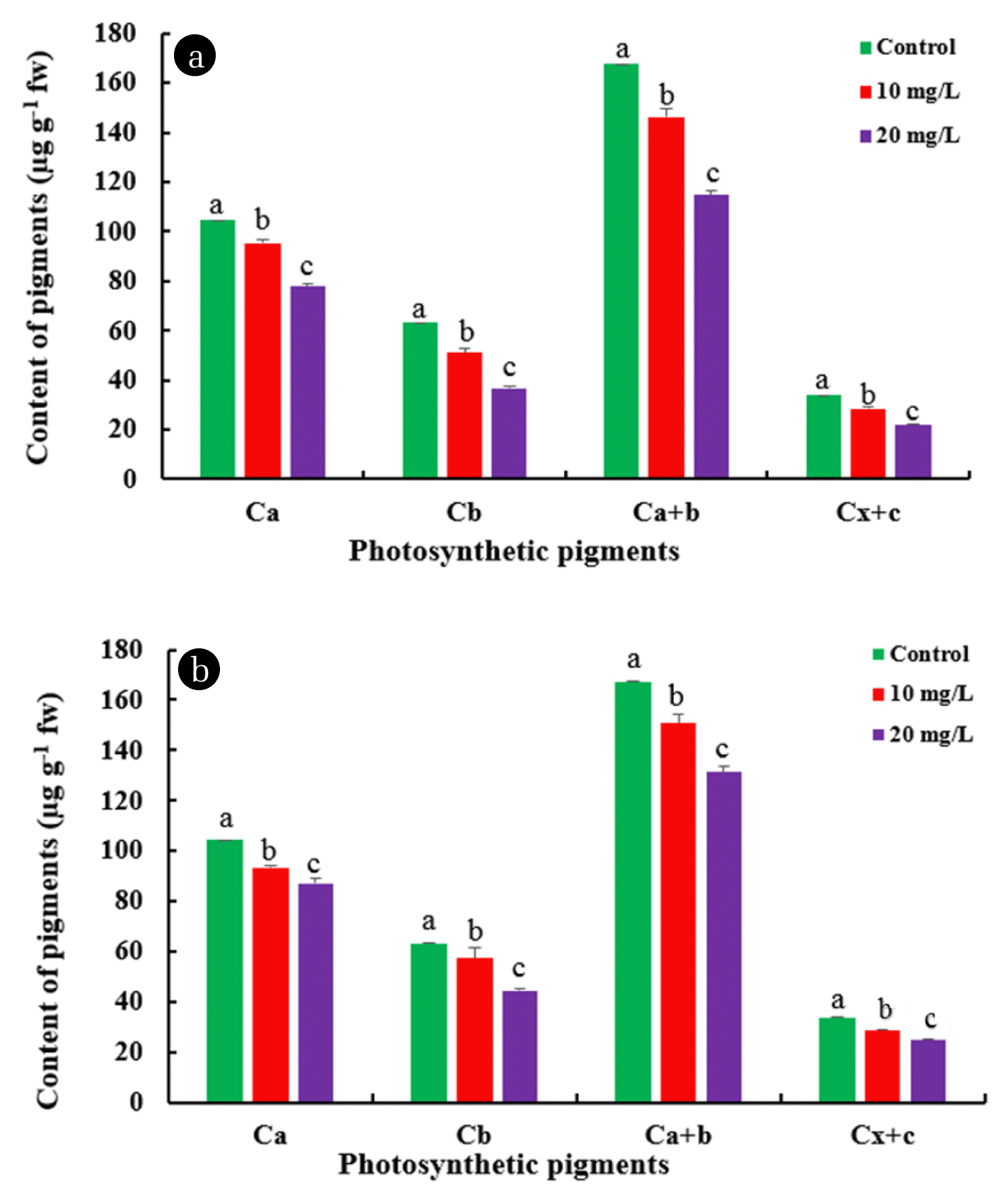
The effect of different concentrations of phenanthrene (a) and pyrene (b) on photosynthetic pigments content of Lemna minor after 10 days of treatment. Different letters indicate significant differences (P < 0.05) according to Duncan Test (mean ± SD, n = 3)
3.3. Accumulation and Degradation of Phenanthrene and Pyrene by L. minor
An HPLC analysis was executed to confirm the entrance and uptake of PAHs in plant tissues. Fig. 3 shows the concentrations of PAHs taken up by L. minor at the end of the 10th day after treatment. Accordingly, the quantities of phenanthrene in the 10-day treatments with 10 and 20 mg L−1 were 0.03 and 0.195 mg g−1 FW, respectively (Fig. 3(a)). On the other hand, the amounts of pyrene in 10 and 20 mg L−1 treatments were 1.237 and 1.66 mg g−1 FW, respectively (Fig. 3(b)). According to the obtained results, the plant’s concentration of phenanthrene and pyrene was significantly increased by the increasing treatment levels from 10 to 20 mg L−1. In addition, the accumulation of pyrene was higher than that of phenanthrene in all treatments (Fig. 3(a), (b)). These results indicated that possibly phenanthrene was partially metabolized by L. minor during the ten-day treatment, but pyrene was accumulated in the plant. In agreement with our data, bioaccumulation of PAHs by various plant species has been widely demonstrated [1, 26, 38]. Houshani and his colleagues [1] showed that phenanthrene could be metabolized in the tissues of the maize plant, but pyrene had more tendency to be accumulated. Similar results were reported from the accumulation of fluorene as a PAH by Salehi-Lisar et al. [38] in sunflower, wheat, and alfalfa plants over time. The water solubility, molecular weight, lipophilic property, and structure of a compound such as PAHs determine its accumulation and degradation by plants [39]. PAHs with higher molecular weights and lower water solubility, such as 4-ring pyrene, are more stable and recalcitrant to be removed than PAHs with lower molecular weight, such as 3-ring phenanthrene [40–42]. This is probably the main reason for the higher bioaccumulation of pyrene compared to phenanthrene.
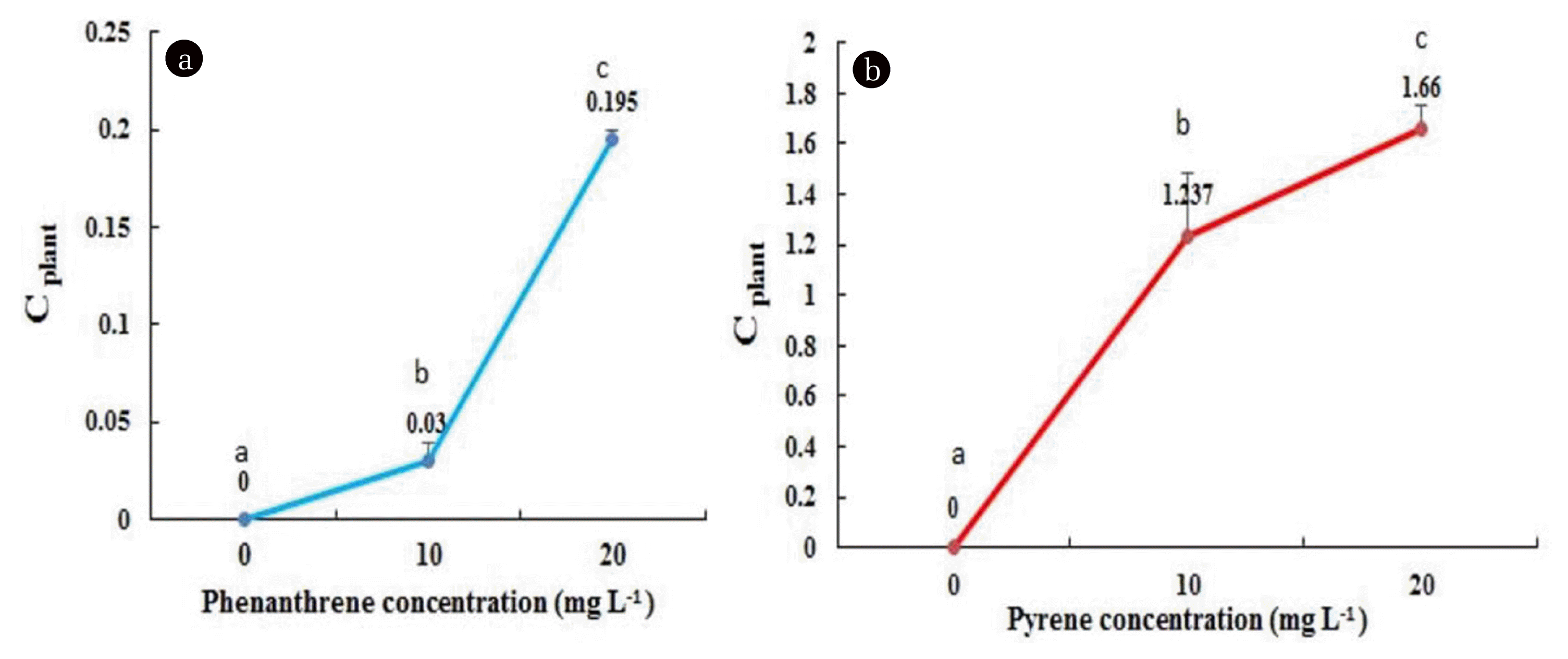
Changes in the concentration of phenanthrene (A) and pyrene (B) (mg g−1 fw) of L. minor plant (C plant) after 10 days of the exposure time. The data are the means of three replications and error bars indicate SD and different letters indicate significant differences (P < 0.05).
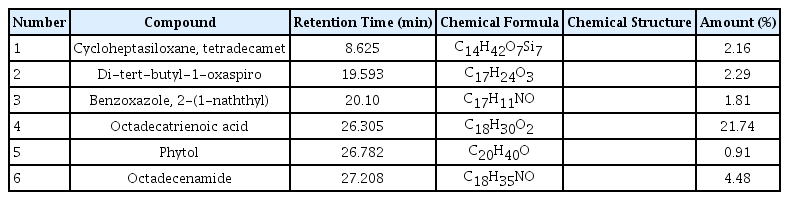
The treated samples with 20 mg L−1 concentrations of PAHs were analyzed using GC–MS to identify the by-products resulting from biodegradation of phenanthrene and pyrene by L. minor. The constituents were identified by matching their spectra with those recorded in the Mass library (Wiley7n and NIST08). Accordingly, six compounds were effectively detected and listed in Table 1 and 2 during phenanthrene and pyrene degradation, respectively. The amount of identified by-products (%) that resulted in phenanthrene biodegradation was higher than the pyrene. In the control sample (free-PAHs solution containing plant), no peaks associated with these compounds were identified using GC–MS. Based on the GC–MS analysis, possible degradation pathways of phenanthrene and pyrene were proposed, as shown in Figs. 4 and 5. In the suggested path, the phenanthrene and pyrene are subjected to aromatic ring cleavage, possibly by oxygenase enzyme system, to form compounds A, B and C. Consequently, these compounds undergo further oxidation for the complete degradation of the benzene ring and the formation of more superficial structures such as single-chain by-products and fatty acids (compounds D, E, and F). In agreement with our results, aromatic ring oxidation by the unique enzyme system, including peroxidase and o-diphenol oxidase in alga and plants, has been previously reported [13, 26, 36, 43, 44]. Patel and Tiwari [36] showed that microalgae Chlorella vulgaris degraded two PAHs (fluoranthene and acenaphthene) and explained the detection of the degradative enzymes and the metabolites resulting from the PAHs biodegradation process. Although, Vafaei et al. [13] reported six intermediated compounds by the cleavage of aromatic dye molecules by peroxidase enzyme in plant Hydrocotyle vulgaris by GC–MS analysis. Overall, the intermediate compounds observed during the phenanthrene and pyrene degradation in L. minor plants verify both PAHs’ biological transformation.

Identified By-products during Biodegradation of Phenanthrene by L. minor
Identified By-products During Biodegradation of Pyrene by L. minor
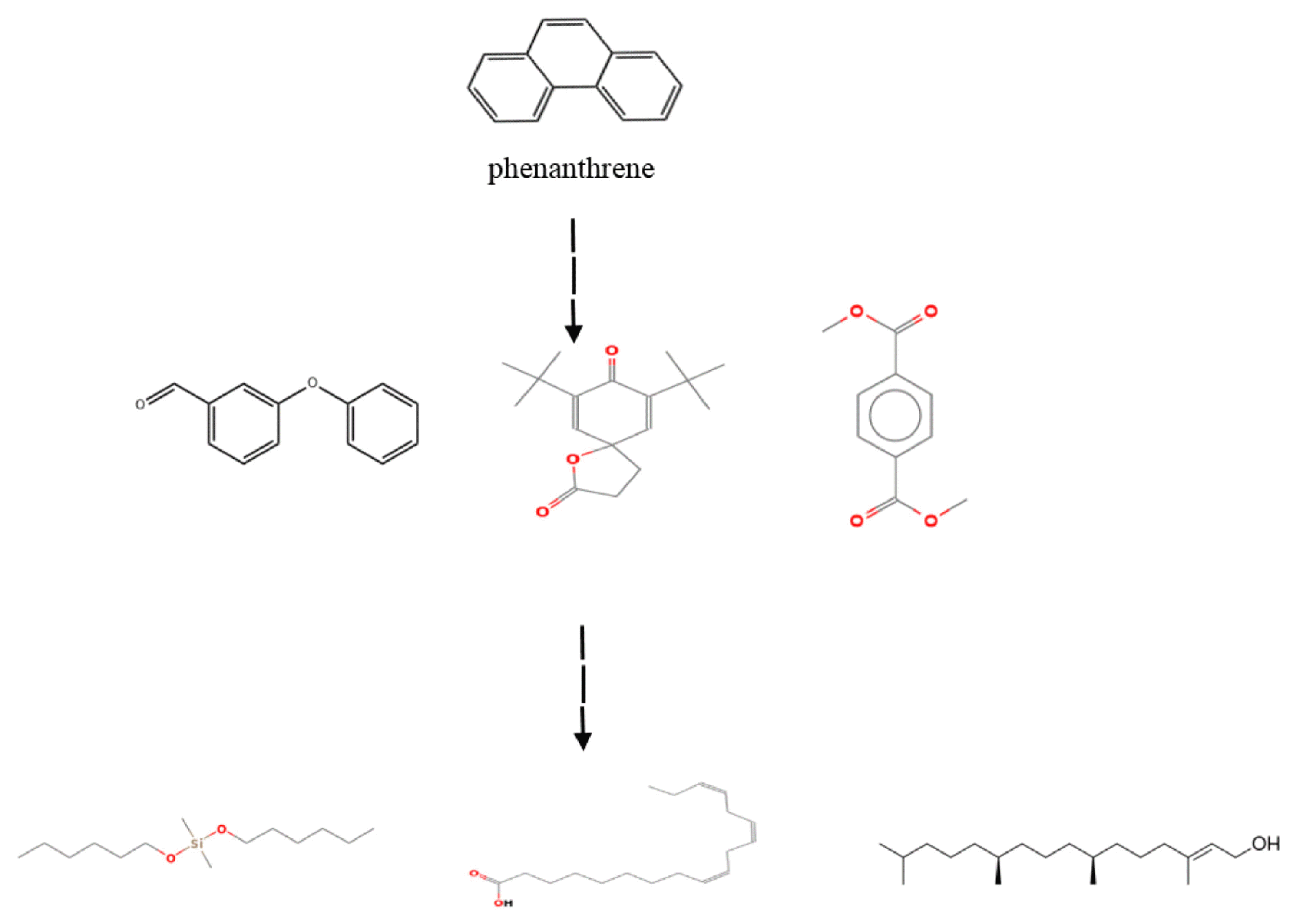
Proposed degradation pathway of phenanthrene by Lemna minor.
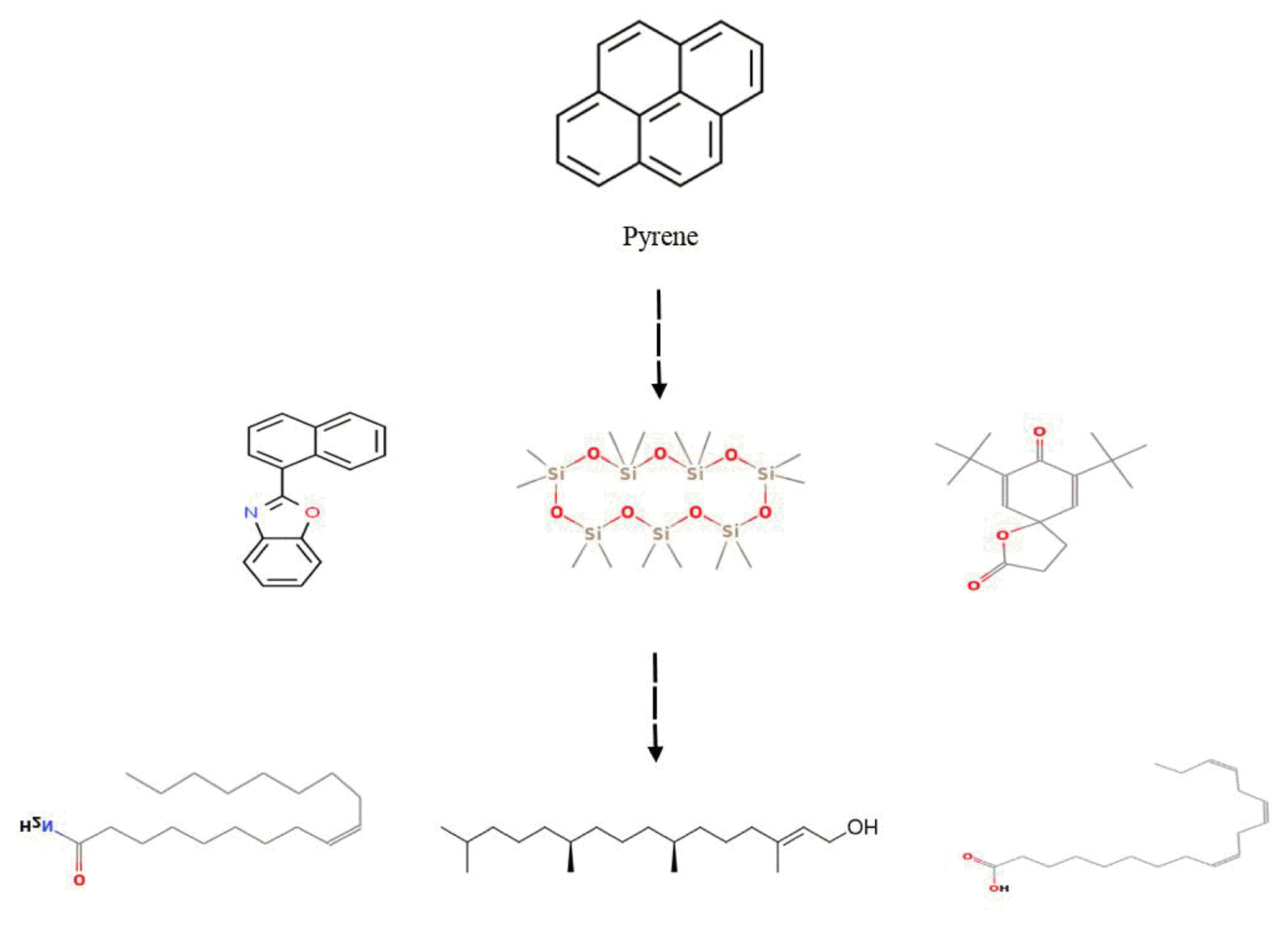
Proposed degradation pathway of pyrene by Lemna minor.
4. Conclusions
In summary, phenanthrene and pyrene displayed significant toxic effects on L. minor, resulting in reducing of growth factors as well as photosynthetic pigment contents in a dose-dependent manner. According to the obtained results of HPLC analysis, phenanthrene and pyrene were found in both treatments after being added into the media for ten days, emphasizing that L. minor efficiently has potential for bioaccumulation of phenanthrene and pyrene. Moreover, the obtained results from GC-MS analyses revealed that L. minor can metabolize both compound after ten days of treatment. Obviously, some biodegraded by-products were identified which were formed by aromatic ring cleavage. Accordingly, possible degradation pathways of phenanthrene and pyrene were proposed. Hereafter, further research is required to thoroughly understand the ways and fate of compounds in plant tissues that could help to enhance the role of L. minor as a vital bioremediation agent against water pollution.
Acknowledgment
Research reported in this publication was supported by Elite Researcher Grant Committee under award number from the National Institute for Medical Research Development (NIMAD), Tehran, Iran. Also, the authors would like to express their thanks to the laboratory staff of the Department of Environmental Health Engineering, Faculty of Health and Health Sciences Research Center for their collaboration, and to the Research Deputy of Mazandaran University of Medical Sciences, Sari, Iran, for the financial support of this study.
Notes
Conflict-of-Interest
The authors declare that they have no conflict of interest.
Author Contributions
M.A.Z. (Professor) was responsible for research design, supervision, validation, article writing, and project management. S.A. (Postdoctoral researcher) was responsible for methodology and conducting experiments, data analysis, and article writing. S.Y.S.L. (Associate Professor) helped in the supply of phenanthrene and pyrene. E.B. (Associate Professor) made contributions to the HPLC and GC-MS analysis. R.T. (Postdoctoral researcher) and N.D. (Master’s student) contributed and critically read the manuscript.