1. Introduction
As highlighted by the Inter-government Panel on Climate Change (IPCC) in 1988, Kyoto Protocol in 1997 [1] and the Paris Agreement in 2015, reducing greenhouse gases is a global concern. The 17th Conference of the Parties (COP) held in 2011 in Durban, South Africa, focused on finding ways to reduce greenhouse gases and the COP adopted the Durban Outcome [2]. According to the outcome, all member countries of UNFCC have to take action towards reducing global warming gases beginning in 2020. Greenhouse gases that trigger global warming include carbon dioxide (CO2), methane (CH4), nitrous oxide (N2O), hydro-fluorocarbons (HFCs), perfluorocarbons (PFCs), and sulfur hexa-fluoride (SF6). The focus of attention has been mainly on reduction of CO2 because this gas forms the majority of emissions [3].
One promising CO2 reduction technology is the use of photosynthetic microorganisms, whose CO2 fixation efficiency can be increased quite simply using a highly concentrated microbial population [4]. The compelling advantages of this technique include the ease of algae in adapting to new environments, simple population growth [5–6], and the abundance of proteins, carbohydrates, and nutrients in the algal biomass that develops during microbial growth through CO2 fixation [4, 6–7]; the resulting biomass can be recovered and utilized as an alternative food or energy source [3, 6–8]. In addition, algal biomass can acts as adsorbent to purify water by toxic metal adsorption [9]. CO2 fixation using photosynthetic microalgae has become a very attractive potential approach to resolve global issues of energy as well as simultaneously increasing the global food supply and water purification.
Spirulina platensis (S. platensis) is a promising microalga for CO2 fixation because of its rapid population growth and production of valuable compounds with a high percentage of proteins, vitamins, and g-linolenic acid [6]. Many studies have addressed the mass production of microalgae in regions that receive high levels of direct sunlight and have a high ambient air temperature. Recently, there has been growing interest in photoautotrophic algal reactors that are engineered for enhanced production rates [3, 6, 10–11].
Many researchers have focused on experimental observations to identify the correlation between key factors and algal cultivation, or to identify optimal algal growth conditions [3, 6, 10–15]. Increased algal production attenuates light capacity, which in turn affects growth rates, making any increased production self-limiting in nature [14]. CO2 and nutrient concentrations are also significant parameters [3, 5, 7–8, 12–20]. In particular, dissolution of CO2 governs pH, which will eventually affect algae levels. However, there is a lack of mechanistic and quantitative analyses of algae biomass concentrations, nutrients, and pH associated with CO2-mixed air flows. All these factors should all be considered to optimize algae production.
In this study, the growth characteristics of S. platensis were investigated using four cylindrical 400-mL photo-bioreactors with CO2-mixed air gas flows. Each reactor was operated under a given flow condition: 3% CO2 at 50 mL/min, 3% CO2 at 150 mL/min, 6% CO2 at 50 mL/min, and 6% CO2 at 150 mL/min. This study further improved the authors’ previous study [21]. The analyses include regression fitting for the algal growth curves, prediction of pH in medium using the reaction equilibrium and kinetics, and stoichiometric assessment of nitrogen (N) limitation.
2. Materials and Methods
2.1. Materials
S. platensis UTEX LB 2340 was obtained from the University of Texas at Austin. The culture medium was alkaline inorganic. The components included NaHCO3 (13.61 g/L), Na2CO3 (4.03 g/L), K2HPO4 (0.50 g/L), NaNO3 (2.50 g/L), K2SO4 (1.00 g/L), NaCl (1.00 g/L), MgSO4·7H2O (0.20 g/L), CaCl2·2H2O (0.04 g/L), PIV metal solution (6 mL/L), Chu micronutrient solution (1 mL/L), and Vitamin B12 (1 mg/1000 mL H2O; 150 mL/L). The PIV metal solution consisted of Na2EDTA (750 mg/L), MnCl2·4H2O (41.2 mg/L), CoCl2·6H2O (2.0 mg/L), Na2MoO4·2H2O (4.0 mg/L), FeCl3·6H2O (97.2 mg/L), and ZnCl2 (5.2 mg/L). The components of Chu micronutrient solution included Na2EDTA (750 mg/L), MnCl2·4H2O (12.8 mg/L), CoCl2·6H2O (20.0 mg/L), Na2MoO4·2H2O (12.8 mg/L), H3BO3 (618.0 mg/L), CuSO4·5H2O (19.6 mg/L), and ZnSO4·7H2O (44.0 mg/L). The medium was utilized for cultivation after being sterilized at 121°C for 15 min in a high-pressure sterilizer. Algae were cultivated in a 250-mL Erlenmeyer flask with stirring at 120 rpm for 12 h under light and 12 h in darkness. The initial pH and temperature of the medium was 9.5 and 30°C, respectively. The light was provided by a 25 mmol/m2-s fluorescent lamp. For algae growth, 400 mL of inoculated medium was prepared using sterilized medium and 0.1 g/L (dry condition) S. platensis. The inoculant medium was injected into the reactor, which had been sterilized beforehand.
2.2. Photoreactors
The cylindrical photoautotrophic bioreactor had a diameter of 4 cm, depth of 65 cm, and volume of 400 mL. The CO2/air mixture was supplied using an air pump and a gas mixer. To prevent contamination by dust or microbes, a 0.2-mm filter was installed at the gas inlet of each reactor. Identical spargers were installed inside the reactor so that microbubbles of CO2 in air could be effectively transferred to the medium. Fluorescent lamps were installed along each side of the reactor. Fig. 1 shows the reactor set-up. The test parameters to investigate the effects of CO2 concentration and air-mixture flow rates were 3% CO2 at 50 mL/min, 3% CO2 at 150 mL/min, 6% CO2 at 50 mL/min, and 6% CO2 at 150 mL/min. The pH of the medium immediately after inoculation was 9.5, and reaction temperature was maintained at 30°C using a water circulator. Light intensity was consistent at 110 mmol/m2-s.
2.3. Analytical Techniques
The amount of microalgal biomass concentration (cell population density) was obtained by measurement of the optical density (O.D.) at 670 nm using a model U-2000 UV spectrophotometer (Hitachi, Chiba, Japan). The dry biomass (= −0.4289×O.D.) was confirmed to have a good linear correlation (r2~99%) with the O.D. The similar analysis to determine cell growth was utilized by Chen et al. and Xie et al. [10–11]. The pH of the medium was measured using a model HI8424 pH meter (Hanna Instruments, Woonsocket, RI, USA). The initial concentrations of N and phosphate (P) in the medium were 305.0 mg/L (NO3-N) and 73.0 mg/L (PO4-P) as measured by ion chromatography (IC) using a DX-120 apparatus (Dionex, Sunnyvale, CA, USA).
2.4. Data Analysis
The exponential and linear regression curves were provided to characterize the dynamic growth curves for each reaction set. The coefficient of determination was found for each curve to assess the fit in the regressions to the corresponding experimental data. Also, Analysis of Variance (ANOVA) was performed with the regression parameters to compare the dynamic algal growth between individual reaction set. If any regression of a data set could not properly characterize a dynamic curve, the data set would undergo a simple treatment (such as geometric averaging) and then conduct the multiple comparison. The regression and the comparative statistical analysis were implemented using MS-Excel and R software.
3. Results and Discussion
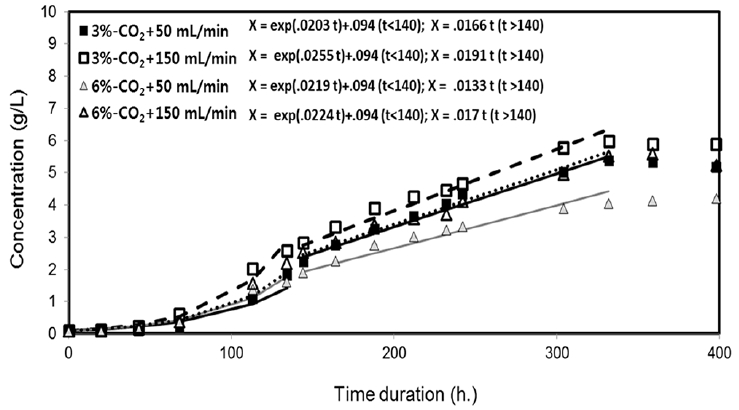
The dynamic cell-growth curves and their regression fits on S. platensis and the dynamic variation of medium pH are shown in Fig. 2. The highest algal biomass concentration and growth curve were obtained for 3% CO2 at 150 mL/min. The 6% CO2 at 150 mL/min condition produced a similar growth pattern, while 6% CO2 at 50 mL/min yielded the lowest growth. Interestingly, both sets with a flow rate of 150 mL/min showed higher S. platensis growth than their counterparts with a flow rate of 50 mL/min. The similar trend of the influences of CO2 contents or aeration flow rates to the algal growth was observed in the previous studies [12–13].
With the respect to the model fits, the early phase of the algal growth in Fig. 2 was characterized by conventional exponential growth until 134 h, in which the increase of the algal population (dX/dt) has normally assumed to have been proportional to the concentration of the algal biomass concentrations (X) [3, 14]. The specific growth rate in the exponential growth can be simply calculated as (ln X2 - ln X1)/(t2 - t1), by applying an ordinary differential equation (ODE) solving technique of the separation of variables. However, a linear increase of biomass concentration predominated in mid-to-final stages of the cultivations (144–332 h). In this study, linear growth rate was described using the model presented by Ogbonna et al. [14]. The model was constructed with the assumption that the growth rate is proportional to the maximum total light energy absorbed in a reactor divided by the liquid volume of the reactor. Here, the constant specific growth rate of the original exponential growth (dX/dt = mX) is modified to be a function that is inversely proportional to the biomass population concentrations X (dX/dt = KaL). It is quite reasonable for the case of absorbed light energy limited by concentrated biomass produced.
The pH of each condition differed significantly according to the CO2 concentration of the gas in the bubbles, regardless of the gas flow rates. At the beginning of the experiment, the pH was 9.5. By 20 h and thereafter, the pH stabilized at 8.8 ± 0.2 for 3% CO2 and 8.5 ± 0.2 for 6% CO2 as similar to the previous studies [3, 10, 12].
Fig. 2 includes the exponential (0–134 h) and the linear regressions for reaction times from 144–332 h. The linear regression equations were included in a range of 0.0138–0.0191 /h, while the constants were different according to the reactor sets. The condition of 3% CO2 at 150 mL/min had the highest linear growth rate.
Here, KaL represents the parameter of the linear growth rate; pH represents the stabilized pH of the solution medium, and Qa denotes the aeration rate. Each growth rate could be well described by linear curves with the coefficients of determination greater than 93%. Based on ANOVA analysis with the F statistic with each linear coefficient, there is a higher significance (p ≈ 0.10) of difference between the individual linear curves with respect to the aeration rates than CO2 levels (p ≈ 0.14). This means the aeration rates can be an effective factor to differentiate the growth curves. Prior to the linear increases until 134 h, the algal growth was assumed to be described by exponential growth curves. The specific growth rates in the exponential growth rates were calculated as 0.02–0.0255/h. However, the specific growth rates had little correlation with the performance of the cultivation of individual sets. This is similar to the results of the linearity shown in many previous studies [3, 6, 13–14]. Ogbonna et al. reported good correlation between the linear growth rates and the final cell concentrations during light-limited batch cultivation of Chlorella pyrenoidosa C-212 and S. platensis M-135 cells, regardless of the type and size of photobioreactor [14].
3.1. pH of the Solution Media
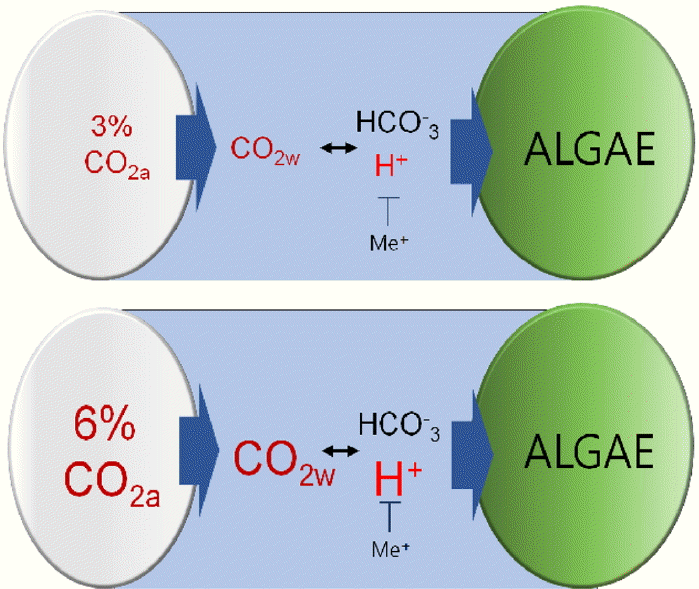
CO2 transferred from the gas to water phases reduces pH by formation of H2CO3, but the H2CO3 is transformed to HCO3− or CO32− at pH 8–9 and pH 10–11, respectively. In this study, additional CO32− or HCO3− ions were initially supplied in the medium with spontaneous dissolution of NaHCO3− and Na2CO3. For that reason, the initial pH was specified as 9.5 by the equilibrium between HCO3− from NaHCO3, and CO32− from Na2CO3. According to the previous studies by James et al., Martis et al. and Stumm and Morgan [7, 15–16], the equilibrium constant ([H+][HCO3−]/[CO32−]) of the reaction between the two ionic compounds HCO3− and CO32− is 10−10.3 at a water temperature of 30°C. This means that at pH 10.3 the two ions have identical concentrations.
The initial pH (~9.5) should rapidly decrease because of the injection of aeration bubbles containing 3% or 6% CO2 gas, which dissolves and is partly transformed to carbonic acid (H2CO3). According to Henry’s constant [18], the dissolved CO2 concentration is linearly correlated with atmospheric CO2. Therefore, 6% CO2 can generate twice the amount of dissolved CO2 in the aqueous medium (≈ 1.1 mM) than 3% CO2 (≈ 0.55 mM) at equilibrium. This is reasonable because other kinds of reactions that feature autotrophic CO2 consumption should be slow at the beginning of the cultivation. H2CO3 is again transformed to HCO3− (> 95%) at pH < 8.5, and HCO3− is the dominant carbonate species at that pH range. In addition, most of the initial CO32− compounds combine with H+ of H2CO3 to form HCO3−. Under the condition of sufficient C as well as the beginning of the reaction, it is reasonable that the gaseous and dissolved CO2 (without H2CO3) should maintain the equilibrium.
Some CO32− and HCO3− ions that exist in the medium may be acidified by H+ in H2CO3, while the dissolved CO2 is converted to HCO3− (through H2CO3). According to the kinetics of the reaction has been described in a previous study [16], more HCO3− is transformed to CO2 and OH− than its reverse conversion (CO2 + OH− → HCO3−) until pH > 8.9 and pH > 8.6, for 3% CO2 and 6% CO2, respectively. At CO2 ≈ 0.55 mM (3% CO2 set) and CO2 ≈ 1.1 mM (6% CO2 set), those transformations stop around pH 8.9 and 8.6, respectively. Termination of pH drop may result not only from activation of the reverse conversion due to increase of dissolved CO2 levels, but also from the charge balance because of the presence of enormous amounts of metal cations (mainly Na+) dissociated from NaHCO3− and Na2CO3, leading to suppress new H+ generation. The mechanism of pH by carbonate and pH is illustrated in Fig. 3.
Typically, algal growth accompanies consumption of CO2, HCO3−, and nutrients, as well as the production of O2 and changes in pH. As for S. platensis, Cornet et al. suggested the following stoichiometric equation [17]:
where CH1.650 O0.531 N0.171 S0.07 P0.006 is a typical element composition of S. platensis. Compared with the old one, there is a different cell composition of S. platensis, recently suggested by da Silva et al. [6].
Eq. (2)-(3) indicates both the metabolic reactions consume H+ and/or produce OH− leading to a potential pH increase, but becomes neutralized by the aforementioned H+ produced by CO2 supplied in the injected aeration bubbles. As a result, rather stable pH levels were maintained over the whole reaction. It has been reported that the optimal pH for biomass production of S. platensis should be around 9.0–9.5, and the amount of biomass decreases as the pH decreases at 30°C if no additional control was made [10, 12]. The present finding of different biomass production with pH supports those of a previous study [13]; the low pH from 6% CO2 should be more disadvantageous for S. platensis growth than that of the 3% CO2 condition. Several studies have described decreased growth of S. platensis by high CO2 in aeration, leading to decreased pH of the medium [3, 12–13]. Park et al. study commented that the high CO2 supply accelerated the growth rates but eventually obtained lower cell biomass concentrations [3].
3.2. Influences of Aeration Rates
There have been few studies on the effects of aeration and gas flow rate on the growth of S. platensis. Tadros showed that algal growth increased with increasing aeration rates in the range of 150–500 mL/min, although growth was deterred at a high aeration rate of 2,000 mL/min [12]. Why aeration affects algal growth remains unclear. The most plausible explanation is the facilitated mass transfer of CO2 from air to algal biomass, but this does not explain the present results, since pH was stable all during growth. Furthermore, pH was more strongly affected by CO2 in the bubbles than the aeration rates. CO2 was not appreciably affected by the rate of aeration. This implies that CO2 mass transfer from the bubbles to the medium should be far greater than CO2 consumption by algae at the beginning of algal growth. Thus, the stable pH is due to the CO2 consumption rate, which remains lower than CO2 mass transfer regardless of CO2 concentration for high algal biomass. The advantages of increased aeration rates may actually result from deformation of microbubbles at high flow rates, leading to improved mass transfer of gases other than CO2, homogeneous mixing of nutrients, or promoted light distribution into individual cells by the mixing.
3.3. Influence of N and P Concentration in Dynamic Cell Growth
Fig. 4 shows NO3-N (Fig. 4(a)) and PO4-P (Fig. 4(b)) concentrations measured in the media at 0, 113, 212, and 398 h. The residual N concentrations at these times were 40–60%, 3–7%, and 0%, respectively, of the initial N concentration for all reaction conditions. In contrast to the huge drop in the residual N concentrations in the middle hours of the cultivation, PO4-P levels gradually decreased, with a loss of only 10–20% at 113 h and ~50% at 212 h. Even at 398 h, about 20–30% of the initial P level remained in the media for all reaction conditions. Like N removal, the conditions of 3% CO2 at 150 mL/min and 6% CO2 at 50 mL/min yielded the lowest and highest P concentrations, respectively, at 113 h.
The residual N and P concentrations in Fig. 4(a)–4(b) were compared using ANOVA according to individual sets. The N concentrations on 113 h and 212 h in Fig. 4(a) were averaged for each set to obtain their geometrical means, and produced F-statistic to assess if there are significance of differences according to aeration rates or CO2 concentrations. Similarly, the P concentrations on 113 h, 212 h and 398 h in Fig. 4(b) were utilized for the ANOVA through the geometrical averaging. As a result, the P concentrations exhibited a marginal significance of difference (p ≈ 0.08) according to aeration rates while other cases (the N concentrations for both sets and the P concentrations with CO2 levels) showed no significance of difference. The P consumption corresponded the trend of cell growth in that both produced the higher significance of differences for the aeration rates than CO2 levels. In contrast, the insignificance of difference in the N concentrations regardless of sets noted that the algal N consumption had little dependence on aeration rates or CO2 levels; another mechanism should be dominant for N consumption.
This study performed a stoichiometric analysis to predict nutrient starvation during growth of S. platensis, based on the presumption that the N consumption was derived by N deficit of nutrient starvation. Nutrient starvation occurs when the available nutrients are much less than the required amounts for algal growth. It is assumed that the required quantities of the nutrients can be determined from a typical elemental composition of S. platensis (CH1.650 O0.531 N0.171 S0.07 P0.006 and CH1.65 O0.5 N0.1), as shown in Eq. (2)-(3), respectively. The weight ratio of N in S. platensis was 8.0% with C:N ratio of 5:1 based on the elemental composition in Eq. (2), while the respective 6% and 8.6:1 of the weight and C:N ratios in Eq. (3). The former one is similar to the result of a previous study measuring C:N ratio of 5.5:1 with N sufficiency, while the latter one is close to the case of N deficiency [13].
Even if the stoichiometric model in this study presumed that the algal growth should terminated when nutrient N is exhausted, decreased N composition inside the algal biomass by down-regulating intensive N metabolism reduced the N consumption as available N was exhausted in the culture media [3, 13]. These studies confirmed that the algal growth we observed could be continued by changing the composition of the algal biomass even after the available N had been exhausted. This explains why the algal growth could be sustained even after the N was almost exhausted in the middle of the culture growth.
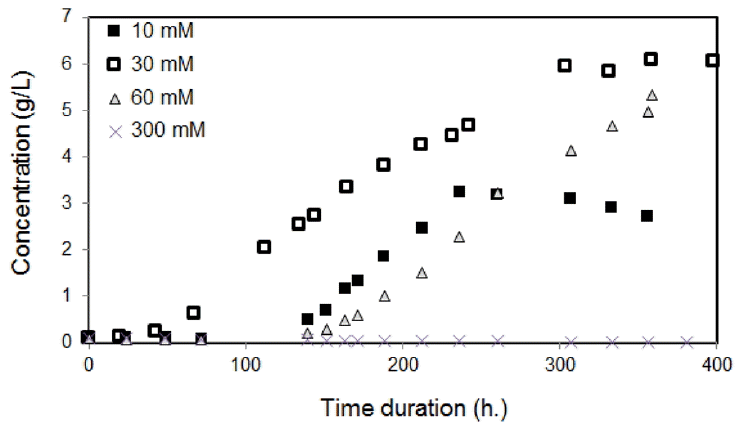
To further investigate the N effects to algal growth, the biomass concentrations were measured according to the various initial NaNO3-N concentrations in the culture medium. Fig. 5 presents the resultant data; the initial N concentrations of 30 mM and 60 mM proved favorable for algal growth. In contrast, 10 mM and 300 mM resulted in premature decline in growth because of lacks of N at the lower concentration and inhibition of growth by the excessively high N. This is entirely consistent with prior findings; even though N is an essential nutrient for algal growth, excessive concentrations restrict algal growth [11, 12]. In the case of P consumption as well as the relevant cell composition in Eq. (2), the quantity of P was relatively sufficient until the end of the growth experiment, P probably must not act as the source of nutrient limitation.
3.4. Performance of the Process
The final biomass concentrations were far higher than in many previous studies with the lab-scaled photoreaction with S. platensis [3, 5, 13, 19]. The biomass concentrations ranged 1–3 g/L which is far lower than this present study with 4–6 g/L. Gordillo et al. [13] obtained low biomass concentrations probably because of the low temperature of the medium (25°C) and use of conventional Plexiglas cylinders as photobioreactor. Kim et al. [5] limited the photoreaction by applying a 16 h light/8 h dark photoperiod. Kumari et al. [19] utilized a special reactor of CO2-agitated sintered disk chromatographic glass bubble column with very high CO2 contents (20–100%). The reactor system may enhance the mass transfer of CO2, but too much high CO2 supply and possible light inhibition in the bubble column may reduce the biomass growth. Park et al. [3] utilized multiple 2 L Erlenmeyer flasks with distant light sources which might be disadvantageous in algal growth, compared to a single reactor with a close light source.
In contrast, the biomass amounts obtained in Chen et al. [10], Cruz-Martínez et al. [20], da Silva et al. [6] and Xie et al. [11] produced comparable to or higher than those in this study. There is no definite clue but they possibly achieved such high biomass concentrations as well as high growth rates by using small diameters of reactors, which could both provide a favorable condition for light energy distribution through the reactor. Especially, da Silva et al. accomplished the biomass production up to 9 g/L with increasing Surface-to-Volume (S/V) ratio in the reactor configuration. This study also incorporated the small diameter of photobioreactor (~4 cm), which maintained high light energy incidence to obtain the high biomass concentrations.
4. Conclusions
Dynamic growth characteristics of S. platensis were investigated under CO2-mixed air flows. Each reactor was operated under a given air flow condition: 3% CO2 at 50 mL/min, 3% CO2 at 150 mL/min, 6% CO2 at 50 mL/min, and 6% CO2 at 150 mL/min. The high gas flow rate of 150 mL/min and the low CO2 content of 3% in aeration yielded high algal growth. The linear growth curves after the exponential growth type properly described the algal growth during active growth, and confirmed the good correlation with the final biomass. A deficit of N might occur after 188 h or more, leading to starvation conditions. Using the stoichiometry between the nutrient amounts and cell elements, it was predicted that depletion of N occurred in the middle of the cultivation but algal growth continued due to change of cell components to reduce N consumption. Based on the pH-carbonate equilibrium and kinetics between dissolved CO2 and HCO3−, the medium pH decreased from 9.5 to 8.7–8.8 (3% CO2) and 8.4–8.5 (6% CO2). The photobioreactors with the small diameter (~4 cm), which allowed favorable light energy distribution, and provision of an acceptable range of CO2 contents (3%) in bubbles as well as proper temperature and medium pH (8.7–9.8) facilitated the accumulation of high biomass concentrations in N-limited conditions.