1. Introduction
Man has revolutionized architectural science for its comfort. Use of marble in structures has long been used by emperors, but with the advent of modernization and excessive use of marble; processing of marble industry has now become a major contributor of toxic heavy metals. It is estimated that 40% of marble industry wastes are drained in farmland, rivers and road sides [1]. Ground water and effluent of marble industries contain high level of lead (Pb), nickel (Ni), arsenic (As), cadmium (Cd), copper (Cu), mercury (Hg) and manganese (Mn), as documented by United States Environmental Protection Agency (USEPA) [2].
Bioremediation is emerging and effective method to remove excessive metals from soil. It is non-hazardous, environmental friendly, cost effective, and non-laborious practice to remove toxic metals [3]. Plant growth promoting rhizobacteria (PGPR) having heavy metal tolerances potential are vital partner of bioremediation [4]. Screening of heavy metal tolerant bacteria has proved beneficial, because they not only alleviate heavy metals toxicity, but also improve plant growth and yield [5]. Recent studies reveal that PGPR having ability to improve plant growth by P-solubilization, nitrogen fixation and phytohormones production are also able to cope with heavy metal stresses [6–7].
Metals found in trace amount like Mn are indispensable micronutrients in metabolic pathways of plants, but the excessive accumulation of these trace elements has deleterious consequences on plants, animals and humans. Heavy metal toxicity may result in several human disorders including damages in blood components and vital organs. It also leads to reduced energy levels and impaired or reduced mental and central nervous function [8]. Beyond certain limit, deposition of these metals becomes toxic to the plants [9].
Mn is micronutrient which is required for plant health and its presence become more important when plants are growing in acidic soil [10]. Excessive accumulation of Mn in low pH soil leads to phytotoxicity of Mn, as it prevent translocation of important elements by blocking root xylem and damaging chlorophyll machinery; hence leading to chlorosis. In these conditions higher oxidized Mn and oxidized phenolics accumulates in leaves and apoplast which result in spotted and necrotic brown leaves [11]. Excessive uses of Mn rich food leads to brain diseases like Manganism and Parkinson disease in human [12].
P-solubilizing soil microorganisms (PSM) release P from organic and inorganic pools of P by mineralization and solubilization and improve crops yields [13]. The affectivity of Bacillus safensis in P-solubilization in sandy fluvo-aquic soils and its impact on growth of wheat has been previously revealed [14]. P-solubilizing Pantoea agglomerans as part of consortium of other PSM was found to increase growth and nutrient uptake in ryegrass [15]. These microorganisms are capable to release various organic acids in soil which are responsible for completing this task [16].
The roles of phytohormones in decreasing heavy metal stresses have been previously documented [17]. The Indole-3-acetic acid (IAA) producing PGPR positively impacted on growth and yields of different plants and cope deleterious effects of heavy metals [18]. IAA has been reported as stress hormone as it enabled plant to cope with metal stresses effectively [19].
Pakistan has agriculture based economy but still 32500 hectares of land is irrigated with industrial toxic waste water, which is fetal for soil health [20]. The Hattar industrial area, Haripur district is situated in northwest of Islamabad at a distance of 60 kilometer. Many industries like pharmaceuticals, heavy electrical engineering, textiles, steel, chemicals, marble tiles and edible oils are working in Hattar. Effluents of these industries accumulate various heavy metals in soil.
Bacteria surviving in marble effluents with high deposition of Mn may act as Mn-tolerant bacteria and prove as an effective tool for reclamation of Mn stressed soil. Such bacteria having plant growth promoting features may be handful as bio-inoculants for crops growing in Mn stressed soil. Some recent researches revealed the capability of Bacillus safensis and Pantoea sp in improving growth and physiology of different plants, either by phosphate solubilization or phytohormone production ([14–15]. However, the role of P. Eucrina and B. safensis as PGPR under heavy metal stress has not been explored so far. The aim of this study was to isolate, characterized and explore Mn tolerant bacteria from soil, affected by effluents of marble industry. The remediation and plant growth promoting ability of P. eucrina and B. safensis, under induced Mn-toxicity and naturally contaminated were determined. The potential of these bacteria in improving growth, and physiology of maize was checked by the bio-inoculation of these bacteria.
2. Materials and Methods
2.1. Isolation and Screening of Bacteria
Twelve soil samples were collected from Hattar industrial area (33.92° N, 72.88° E) in the month of March and stored at 4°C. Collections were made from the land receiving marble effluent and industrial deposits. The temperature, humidity and rain fall of the area were 32 °C, 51% and 68.6 mm, respectively at the time of collection. Samples were collected from three different zones i.e from surface, at the depth of 10–15 cm and from rhizosphere of Conyza canadensis, Euphorbia sp, Parthenium hysterophorus and Amaranthus viridis growing in polluted soil.
Several bacteria were isolated from these samples. Among these, four different isolates were screened out on the basis of Mn tolerance, P-solublization and (Table S1). Pure colonies of these isolates were sent for sequencing to Macrogen, Inc., Seoul, Korea. On the basis of 16S rRNA sequencing results, isolates were identified as Rothia sp, Methylobacterium sp, Pantoea eucrina and B. safensis. Nucleotide sequences were submitted to NCBI and accession numbers MH605007 for Rothia sp, MH603683 for Methylobacterium sp, MH 603867 for P. eucrina and MH603952 for B. safensis were obtained. Two isolates viz. P. eucrina and B. safensis were selected for bio-inoculation on maize under greenhouse condition.
2.1.1. Mn tolerance of bacteria
Mn tolerance of bacterial isolates was determined by the method of [21]. Stock solution of MnSO4 (1000 ppm) was prepared, and from this stock solution working solutions of 1500 ppm, 1750 ppm and 2000 ppm were made. Luria Bertani (LB) agar media was prepared in these working solutions and different sets of petri plates containing different ppm concentration of MnSO4 were made. Decimal concentration 109, 107 and 105 of bacterial cultures (30 ml) were spread over the plates. Plates were incubated at 28°C for 24–48 h and colony forming unit (cfu) were calculated according to the formula given by [22]. The experiment was reiterated in triplicate and agar plates without heavy metal salts were taken as negative controls.
2.1.2. Phosphate solubilization ability
All bacterial isolates were checked for phosphate solubilizing character on Pikovskaya’s agar medium and National Botanical Research Institute Phosphate media (NBRIP) media as expressed by [23]. NBRIP supplemented with different ingredients containing glucose, 10 g; Ca3 (PO4)2, 5 g; MgCl2·6H2O, 5 g; MgSO4.7H2O, 0.25 g; KCl, 0.2 g and (NH4)2SO4, 0.1 g; Fe3 PO4 (2.0 g L−1), Aluminium phosphate AlPO4; 5g L−1 and soy lecithin (15.0 g L−1). Fe3PO4 and AlPO4 were supplemented because Mn rich soil are acidic and accumulate these forms of phosphate. Autoclaved culture media (25 ml) was poured in each petri plate and 10 μL bacterial suspensions were placed at the center. For assessing solublization zone and colony diameters, plates were incubated at 28 °C for 15 days and results were interpreted on the bases of different sizes of both.
2.1.3. Organic acid production of PGPR
Organic acid production and identification were determined by the method explain by [24]. 1 ml aliquots from 24 h old bacterial culture were centrifuged at 3500 × g. Media was filtered through 0.45 millipore filter (Greiner, NC, USA) and centrifuged again. Supernatant was collected in a 96-well plate (Greiner, NC, USA). Organic acids were analyzed by HPLC (Shimadzu, C-R4A Chromatopac; SCL-6B system controller) equipped with a binary pump, a UV and RI detector, and a HPX-87H column (Bio-Rad, CA, USA). H2SO4 (10m mM) was used as mobile phase and flow rate was maintained at 0.5 ml min−1, column compartment was 85 °C, while RI detector was operated at 55 °C. External calibration was used to identify and quantify by different organic acid standards (formic, isobutyric, acetic, lactic, butyric, malic, propionic, gluconic). Identification of different organic acids was made by comparing retention times and the peak areas of chromatograms of samples with their subsequent standards.
2.1.4. Quantification of indole acetic acid (IAA) on Salkowski reagent and high pressure liquid chromatography (HPLC)
The bacterial culture was grown in a 250 ml Erlenmeyer flask containing 50 ml of nutrient broth for 48–72 hours at 28°C. The development of pink color indicated IAA production. The flasks were inoculated with different bacterial strains, respectively. The fully cultured culture was centrifuged at 3000 × g for 30 minutes. Two drops of orthophosphoric acid and 4 ml of Salkowski reagent (50 ml, 35% perchloric acid, 1 ml, 0.5 M FeCl3 solution) were mixed with the supernatant (2 ml). The development of pink indicates the production of IAA. The optical density (OD) of each sample was determined by spectrophotometer at 530 nm. The quantification of IAA was made by standard IAA curve.
For the confirmation of IAA production bacterial cultures were also subjected to high pressure liquid chromatography (HPLC). Yeast Manitol Agar (YMA) broth media (10 mg/100 ml) was inoculated with 24 h old bacterial cultures and placed on a shaker (ECELLA E24, USA) at 100 × g for 5 d. Bacterial cells were harvested by centrifugation at 10,000 × g for 15 min at 4°C and supernatant was collected as described by [25]. The sample was analyzed on HPLC (Shimadzu, C-R4A Chromatopac; SCL-6B system controller) equipped with UV detector and C-18 column (39 × 300 mm) for identification of hormones. Prior to injection in HPLC column, the sample (100 μl) was filtered through 0.45 millipore filter. Methanol, acetic acid and water (29: 1: 70) were used at flow rate (0.5 ml/min) for an average run of 15 min/sample. Pure IAA (sigma, USA) was used as standard and IAA was identified on the basis of retention time and peak area. The detection of IAA was made at 280 nm.
2.2. Green House Experiment
For bio-inoculation experiments, maize (Zea mays L.) cv. Azam was obtained from Baffa Research Station, Mansehra. Garden soil and soil naturally polluted with marble effluents were collected from Hazara University and Hattar industrial area, respectively. Soils were autoclaved and poured into pots with 3:1 ratio of soil and sand. Seventeen treatments were made and each treatment had five replicates. Different concentrations of MnSO4 were provided with irrigation water before sowing. Recommended dose of chemical fertilizers (0.082 g/10kg DAP and 0.035g /10kg of urea) was added to all the treatments except control and pots containing naturally contaminated soil at the time of sowing. In each pot 7–10 seeds were sown and thinned to three plants per pot.
P. eucrina and B. safensis were inoculated alone as well as in consortium with three different concentrations (1500 ppm, 1750 ppm and 2000 ppm) of MnSO4. Controlled plants had no inoculation but were stressed with three concentrations of MnSO4. P. eucrina and B. safensis were also inoculated alone and in consortium in naturally polluted soil collected from Hattar. Control plants were also grown in same soil but they did not receive any bio-inoculant.
Seeds were sterilized according to the method of [26]. Sterilized seeds of maize were soaked for 3 h in microbial culture of P. eucrina and B. safensis having 107cell/ ml. Seeds were shade dried for 45 min and sown in pots. For second set pot experiment seeds were soaked in microbial culture along with different concentrations (1500 ppm, 1750 ppm and 2000 ppm) of MnSO4 for 3 h and then sown after shade drying. Control seeds were soaked in autoclaved distilled water before sowing. Plants were harvested at two-leaf stage, after 60 days of sowing for growth and physiological parameters.
2.2.1. Density of bacteria in rhizospheric soil
P. eucrina and B. safensis were re isolated at 60 DAS and 155 DAS from treated soil by serial dilution preparation [27].
2.2.2. Heavy metal analysis of soil
Soil samples affected with marble effluents were analyzed for different heavy metals by Ammonium Bicarbonate-DTPA method described by [28]. 1.97 g of 0.005 M diethylenetriamine penta acetate (DTPA) was mixed with 79 g of ammonium bicarbonate (NH4HCO3) and 800 ml of distilled water. Volume of the solution was raised up to 1 L using distilled water, and the pH was adjusted to 7.6 by using ammonium hydroxide.
Air-dried soil (10 g) was added in 10 ml of above solution (extraction solution) and shaken for 15 minutes in a shaker at 180 cycles/minute. The extract was filtered through Whatman No. 42 filter paper. The filtrate was analyzed for the presence of Mn through atomic absorption spectrophotometer (Shimadzu AA-700).
2.2.4. Biological concentration factor (BCF), biological accumulation coefficient (BAC), and translocation factor (TF)
2.2.5. Chlorophyll, soluble sugar and proline contents of leaves
2.2.6. Superoxide dismutase assay
The SOD activity was determined by the method of [35], which determines the inhibition of photochemical reduction of nitrotetrazolium (NBT) as measure of SOD activity. A single unit of SOD defined as the amount of enzyme, which reduced the absorbance reading by 50% as compared to the control (lacking enzyme). For the quantification of peroxidase, method devised by [36] was applied. Peroxidase activity was recorded by the change in absorbance at 485 nm at 1 min and after 3 minutes with spectrophotometer and articulated as ΔOD485nm/min mg proteins.
2.3. Statistical Analysis
Treatments were statistically analyzed by Statistix 8.1 software for two ways of ANOVA. All the statistical analyses were carried out on the raw data except antibiotic results, which was transformed before analysis. Completely randomized design (CRD) was applied. Duncan’s multiple range test was performed to determine Analysis of variance (ANOVA) and least significant difference (LSD) among treatment at P≤ 0.05.
3. Results and Discussion
3.1. Mn tolerance of PGPR
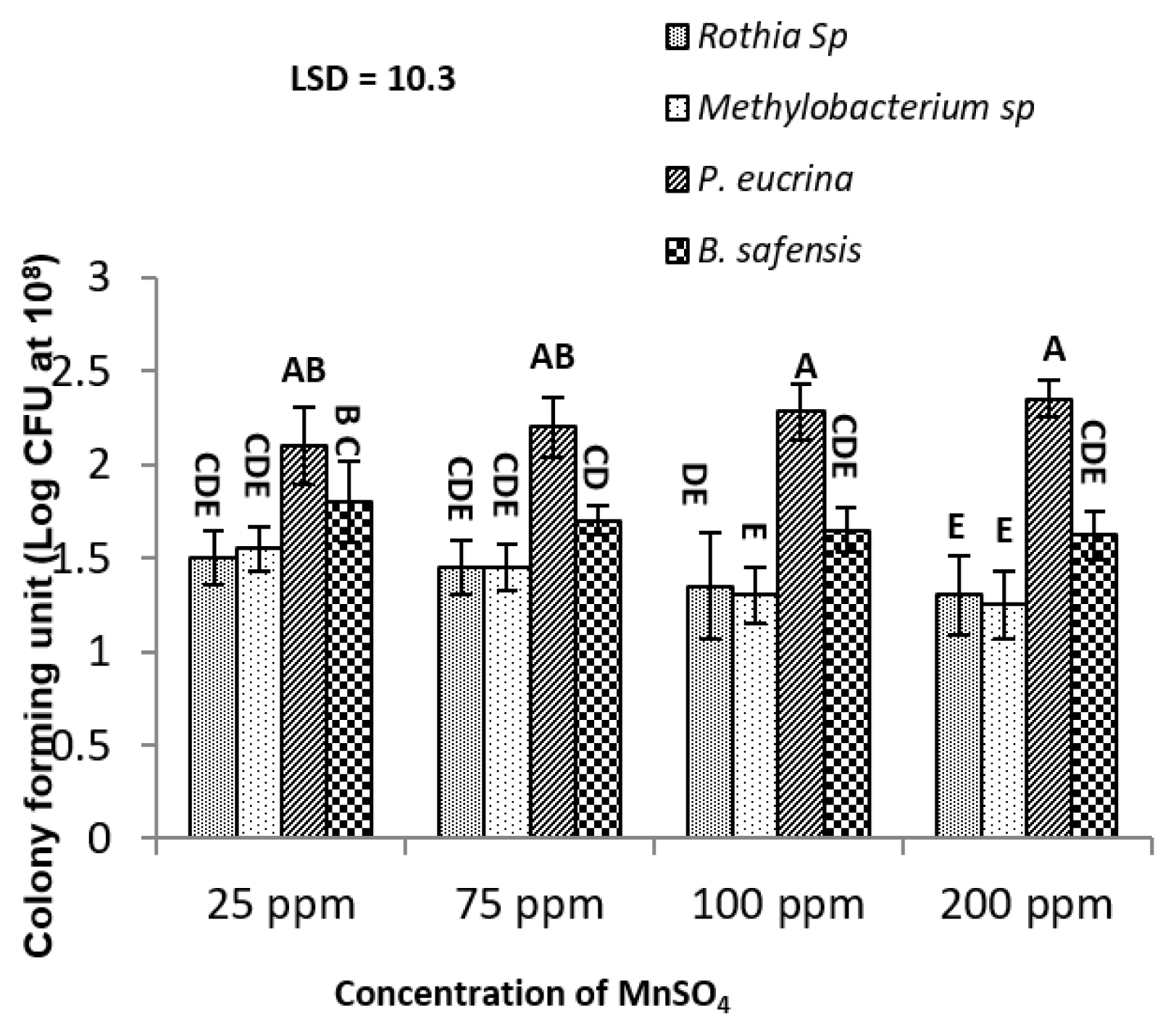
All bacterial strains were tolerant against Mn at 1500 ppm, 1750 ppm and 2000 ppm. P. eucrina and B. safensis were more tolerant than Methylobacterium sp and Rothia sp (Fig. 1). At 2000 ppm, P. eucrina exhibited highest (2.45×106 CFU), while B. safensis 2.2 ×106 CFU at same concentration of Mn. Results indicates that P. eucrina and B. safensis had 96% greater resistance to Mn than Methylobacterium sp and Rothia sp. Similar trend was observed at 108 decimal dilutions (Fig. 2).
3.1.1. Phosphate solublization potential
Phosphate solublization potential measured by inhibition zone of P. eucrina and B. safensis was greater than Methylobacterium sp and Rothia sp (Fig. S1). P. eucrina had 1.6 mm inhibition zone, while Methylobacterium sp had smallest (1.05 mm) zone of inhibition. P. eucrina and B. safensis had 43% and 52% greater zone than Methylobacterium sp.
3.1.2. Organic acids production
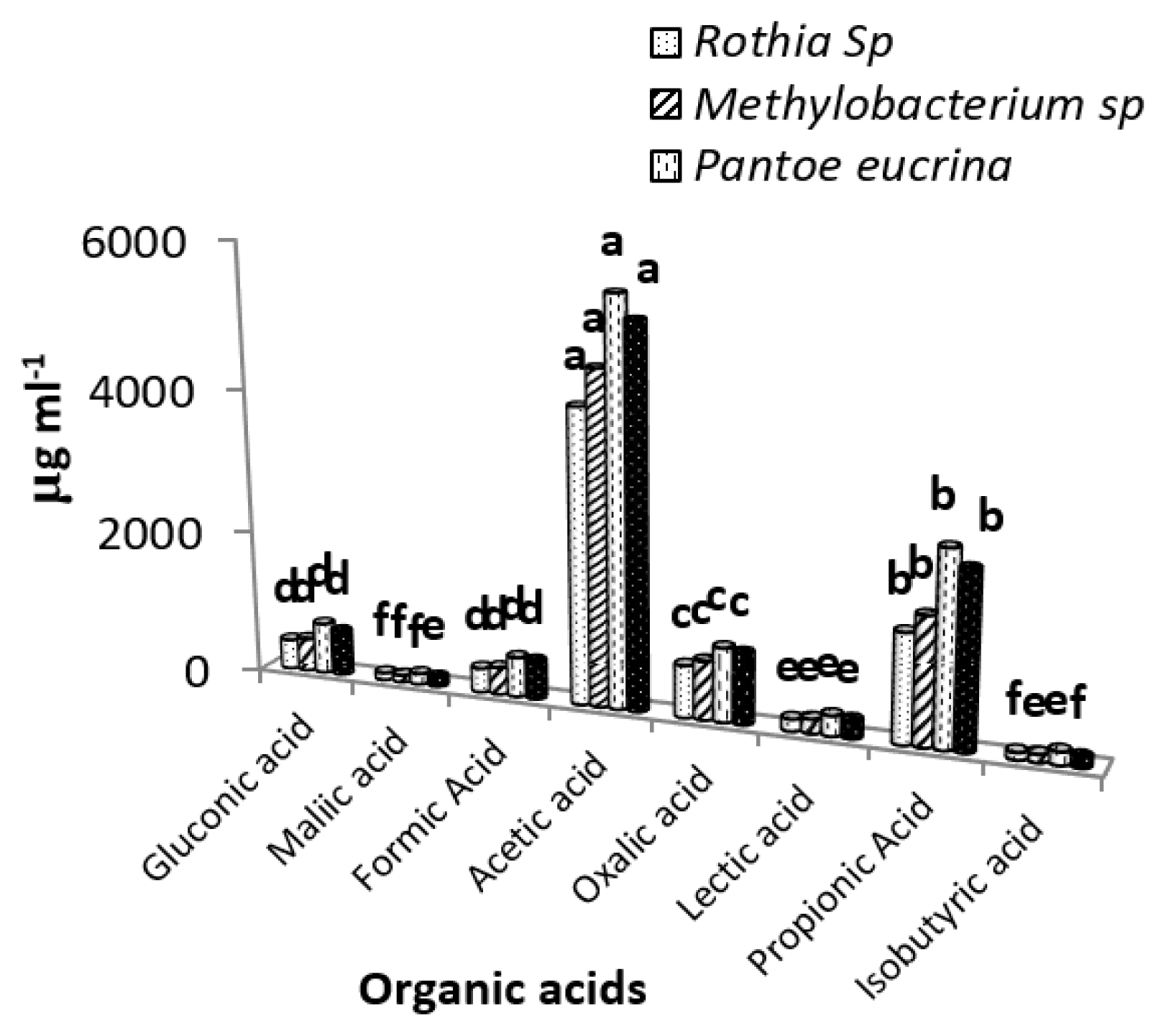
P. eucrina, B. safensis, Methylobacterium sp and Rothia sp produced different concentrations of gluconic acid, maliac acid, acetic acid, formic acid, oxalic acid, lectic acid, propinic acid and isobutyric acid (Fig. 4). Among four PGPR organic acids production was higher in P. eucrina and lowest in Rothia sp. Formic acid and gluconic acid production was 62% and 70% higher in P. eucrina as compare to Rothia sp. P. eucrina produced 50%, 37%, 41%, 68%, 71% and 80% higher maliac acid, acetic acid, oxalic acid, lectic acid, propinic acid and isobutyric acid than Rothia sp, respectively. Similarly, B. safensis produced 51%, 27%, 58%, 28%, 35%, 51%, 60% and 40% higher gluconic acid, maliac acid, formic acid, acetic acid, oxalic acid, lectic acid, propinic acid and isobutyric acid than Rothia sp, respectively.
3.1.3. Indole-3-acetic acid potential
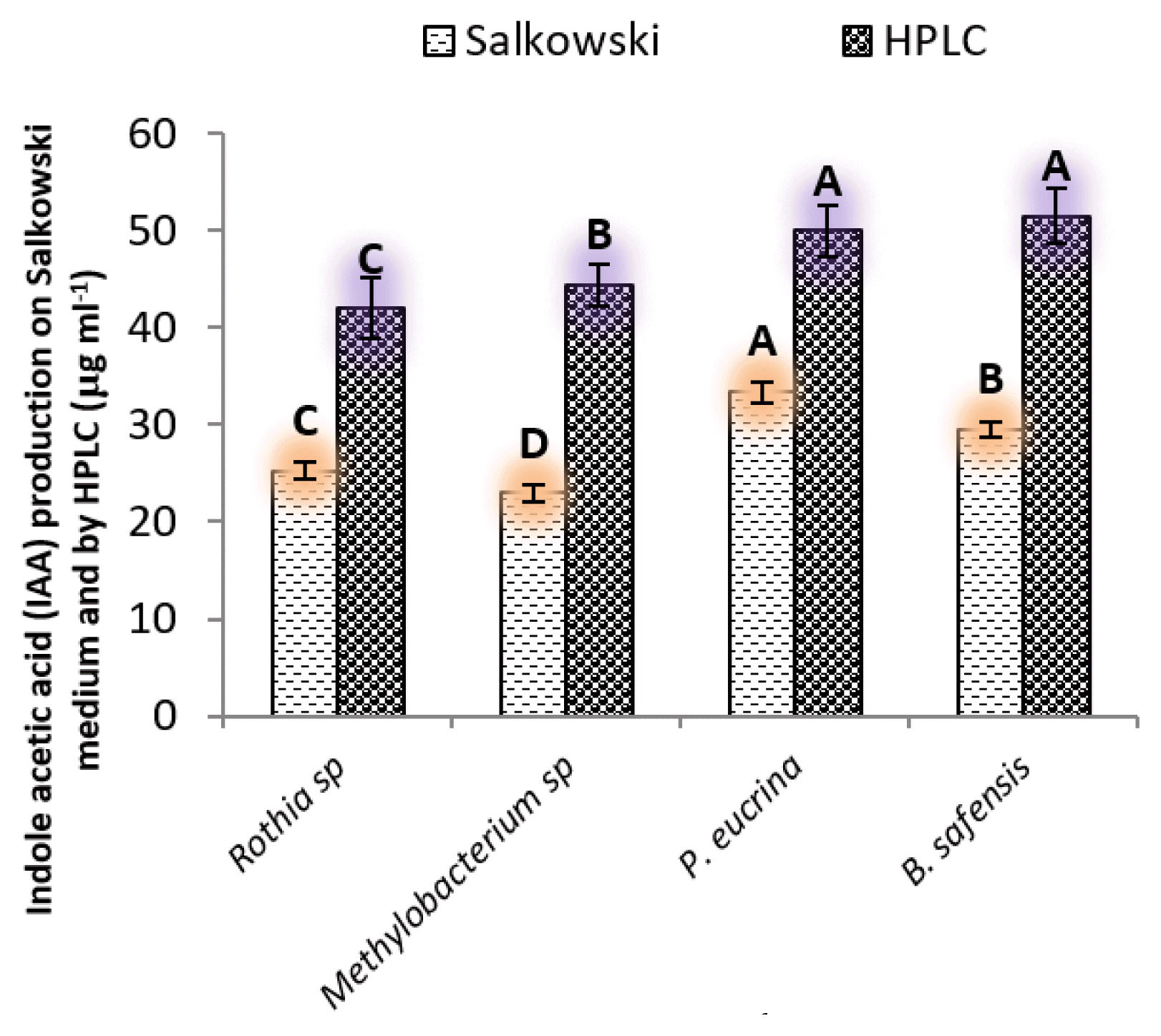
Indole-3-acetic acid production was observed in all strains (Fig. 3). Among bacterial strains, B. safensis produced the highest amount (30.5 and 52 μg ml−1) on Salkowski media and HPLC, respectively. Rothia sp produced lowest (21 and 42 μg ml−1). P. eucrina and B. safensis showed 43% and 56% greater IAA than Methylobacterium sp. B. safensis produced 37% greater IAA than P. eucrina.
3.2. Bacterial Density of Inoculated Bacteria in Soil
A gradual decline in survival of P. eucrina and B. safensis was observed at 60 DAS as compared to CFU at 0 DAS (8.3 × 109) and with increase in Mn concentrations (Fig. S2). However, CFU in single inoculation of P. eucrina and B. safensis was increased by 4–6% as compared to CFU recorded at 0 DAS. Highest increases (8–12%) in CFU were observed in single inoculation of P. eucrina and B. safensis inoculated in naturally polluted soil. Both isolates had lowest CFU in soil received 2000 ppm of Mn.
3.2.1. Effects of PGPR inoculation on bioremediation of Mn
Affectivity of P. eucrina to remediate Mn from soil was greater than B. safensis (Table 1 & 2). P. eucrina and B. safensis decreased 28% and 24% Mn in the soil which received 1500 ppm–2000 ppm of Mn stress with RDF as compared to un-inoculated soil with 1500 ppm–2000 ppm of Mn stress and RDF. In similar cases, decrease in Mn was 30% when consortium was applied with different concentrations of Mn and RDF.
In naturally polluted soil, inoculation of P. eucrina decreased 29% Mn over naturally polluted control soil B. safensis decreased 27% of Mn over control while consortium of P. eucrina and B. safensis decreased 31% Mn in soil.
3.2.2. Mn accumulation in plant parts
Maize roots and leaves accumulated 23–28% and 25% higher Mn when P. eucrina was applied with different concentration of Mn and RDF as compare to un-inoculated control with different concentrations of Mn and RDF (Table 1). B. safensis inoculation with different concentrations of MN and RDF exhibited 17% higher Mn in roots and 20–24% higher Mn in leaves. Accumulation of Mn in roots and leaves were 28–36% higher in consortium treatment over un-inoculated control.
In plants growing in soil contaminated with marble industry, application of P. eucrina increased 28% Mn in roots and 37% in leaves while B. safensis showed 22% and 33% greater Mn in plant roots and leaves over control. Consortium of B. safensis and P. eucrina accumulated 33% and 40% greater Mn in roots and leaves of maize plant.
BAC and BCF of Mn was highest (0.8) and (0.45) when P. eucrina and B. safensis were inoculated with 2000 ppm of Mn and RDF (Table 2). TF of Mn was highest (2.3) in single inoculation of P. eucrina and B. safensis as well as in consortium. In naturally polluted soil increase in BAC and TF was significant (13–15%) in inoculation treatments over control.
3.2.3. Effects of PGPR inoculation on maize growth
Increase in plant height and fresh weight measured at 60 DAS was 20% and 18% over control following the single inoculation of B. safensis and P. eucrina (Table 3). Co-inoculation of PGPR comprising B. safensis and P. eucrina increased plant height by 26% over control. Plant height and fresh weight were 25–30% higher in inoculation treatments as compared to the applications of B. safensis and P. eucrina with different concentration of Mn + RDF. Plants growing in soil contaminated with marble industry had 11–12% higher plant height and 20% higher fresh weight in single inoculation of P. eucrina and B. safensis over control. Consortium of B. safensis and P. eucrina exhibited 64% and 32% greater plant height and fresh weight over control, respectively.
Single inoculation of P. eucrina and B. safensis increased shoot dry weight by 11–13% over control. Consortium comprising P. eucrina and B. safensis had 21% higher shoot dry weight. Under induced Mn stress, consortium application enhanced 14% dry weight. In plants growing in naturally contaminated soil of marble industry, single inoculation of P. eucrina and B. safensis as well as consortium increased dry weight by 55–60% over control.
B. safensiss and P.eucrina inoculation increased root length and dry weight by 11% over control (Table 3). Root length and fresh weight of maize plants were 15% higher over control when PGPR consortium was applied. Under induced Mn stress increased in root length and root fresh weight was 14–18% higher as compare to un-inoculated plant receiving same concentration of heavy metal and RDF. Maize plants grown in naturally contaminated soil had 52% and 34% greater root length and fresh weight over control when inoculated with P. eucrina or B. safensis. Similarly, 52% and 43% higher root length and root fresh weight were observed in plants inoculated with Consortium.
3.2.4. Effects of PGPR inoculation on physiology of maize
The percentage of increase in soluble sugar content was 33% and 16% over control in single inoculation of P. eucrina and B. safensis, respectively (Table 4). Consortium increased 40% sugar content in maize leaves over control. P. eucrina increased soluble sugar content by 60–65% in the presence of different concentration of Mn + RDF as compared to control. B. safeness in the presence of Mn + RDF showed 20–35% greater sugar content over control. The consortium application with Mn + RDF increased 65–75% soluble sugar as compare to control. Maize plants grown in naturally contaminated soil had 20% and 22% greater soluble sugar over control when inoculated with P. eucrina and B. safensis, respectively while consortium exhibited 31% higher sugar contents.
Proline content was 12% higher in single inoculation of P. eucrina and B. safensis. This increase was 26% over control in consortium treatment. P. eucrina with different concentration of Mn + RDF increased 30–48% proline over control. B. safensis showed 36–40% increase proline content in leaves in the presence of Mn + RDF. Consortium with 1500 ppm, 1750 ppm and 2000 ppm of Mn + RDF induced 52% increase in proline content. Maize plants grown in naturally contaminated soil had 24% greater proline over control when inoculated with P. eucrina or B. safensis while consortium exhibited 28% higher sugar contents
The SOD and POD activities were 21% and 24% higher than control following the inoculation of P.eucrina. B.safensis increased SOD and POD by 21% and 16%, respectively while accumulation of SOD and POD were 26% and 32% in consortium treatment. Application of P. eucrina with different concentration of Mn and RDF increased 39–73% and 44–68% SOD and POD over control while B. safensis with different concentration of Mn and RDF increased SOD and POD by 40–56%. Consortium application with different concentrations of Mn and RDF increased 69–100% SOD and 52–80% POD over control. Maize plants grown in naturally contaminated soil and inoculated with P. eucrina had 29% and 19% greater SOD and POD over control while B. safensis exhibited 27% and 16% higher SOD and POD. Consortium of B. safensis and P. eucrina accumulated 32% higher SOD and 22% higher POD in leaves.
4. Discussion
4.1. Isolation of Heavy Metal Tolerant Bacteria
Rothia sp, P. eucrina, Methylobacterium sp and B. safensis were isolated from the effluent of marble industry, having high deposition of Mn. Marble industry effluents have drastic effects on microbial biodiversity and only few bacterial species can survive in high Mn Stress [37]. Soil health and growth indices of plants are also affected by marble effluents as studied in tomato (Lycopersicon esculentum) [38]. Pantoea sp commonly colonized soil as well as endophytes in different plant species including onion, orchid, eucalyptus and brassica [39–40]. Soil contaminated with heavy metals are reported to harbor different Pantoea sp [41–42].
4.1.1. Solublization of phosphorus by bacteria
All isolates were competent to solublize phosphorus (P) and produce IAA. Since IAA production and P-solublization is peculiarity of PGPR and it has been revealed in many researches [43]. It has been demonstrated that under heavy metals stress, B. Safensis compensate deleterious effects by high solublization of phosphorus and IAA production. B. Safensis was found more effective than other isolates under Mn stress, because of its spore producing ability [44].
4.1.2. Mn toxicity in soil
Mn being trace element exists in different oxidative states and it becomes available to plants at low pH [45]. Bacterial community is capable of producing different organic acids, which help to solublise soil P, decrease soil pH and enable plants to survive at high concentration of Mn. Our results also confirm that all the strains were P-solubliser and capable of producing different organic acids. Decrease in Mn contents is correlated with production of higher oxalic acid which is considered as good Mn solublizer [46].
All the bacterial isolates were tolerant to different concentrations of Mn (1500 ppm, 1750 ppm and 2000 ppm) as determined by CFU. Mn tolerant bacteria Serratia marcescent and Lysinibacillus sp were isolated from Mn stressed soil [47–48]. It was reported previously that survival of Bacillus thuringiensis at 4000 mg Kg−1 in manganese ore having PGPR properties [49]. Different studies prove that methylotrophic bacteria are resistant to high concentrations of heavy metals [50].
4.2. Occurrence of Mn in Maize
Translocation of Mn from soil to roots and from roots to leaves was increased following the inoculation of PGPR. This was also evident by BAC, BCF and TF. Decrease in soil Mn content is attributed with better solublization of phosphorus and increased production of organic acids ([13, 51, 52]; thereby increasing solublization of Mn [53]. Most of bacterial species are good Mn solublizers at higher or neutral pH, but data about Mn solublizers and Mn oxidizing bacteria in acidic soil is limited [54]. It was found that Mesorhizobium australicum had potential to oxidize Mn at low pH [53]. Application of Pantoea sp and Bacillus increased translocation and accumulation of Mn in plant parts of strawberry [55].
4.2.1. Recovery of inoculated bacteria from soil
Result indicates that CFU of inoculated P. eucrina and B. safensis decreased with increasing Mn concentrations when Mn was induced at 60 DAS. However, increases in CFU of both inoculated bacteria were observed when they were applied in single inoculation to control soil as well as to naturally polluted soil. Various factors including inoculum density, inoculation methods, soil humidity, pH, temperature and host plant physiology determines survival of inoculated bacteria [56]. According to [57] root exudate, plant age and genotype also effect survival capabilities of bacteria in soil and these may be causes of variation in CFU in different treatments. Greater decreases in CFU of P. eucrina and B. safensis in the soil treated with different concentrations of Mn may be attributed to increase in proliferation of hazardous ions in the soil.
4.2.2. Improvement in growth indices of maize due to inoculation under Mn stress
Marked increases were observed in term of plant height, shoot and root fresh, and dry weight of maize. Increase in growth responses following the inoculation of PGPR is mediated by better acquisition of P and IAA production [58]. Same mechanism was described under drought stress where Pantoea alhagi was found effective to increase growth of wheat [59]. Bio-inoculation of Pantoea sp on Brassica napus has been documented to improve shoot and root biomass [40]. Affectivity of B. safensis was also experimented for growth responses in maize and soybean [60, 61].
4.2.3. Improvement in physiology of maize due to inoculation under Mn stress
Increases in sugar, proline and antioxidative enzyme activities were observed when plants were treated with PGPR and these increases were further augmented in consortium treatment. Sugar, proline and antioxidants are the major osmoprotectants and indicators of stresses [58]. Onset of abiotic stress triggers accumulation of different types of sugars (mannitol, trehalose and galactinol) in plants [62]. Similarly, imino acid proline accumulation is primary indicator to abiotic stresses including heavy metal stresses [63].
Under heavy metal stress, antioxidant enzymes are activated for quenching reactive oxygen species (ROS) which are produced in abundant and damage macromolecules [64]. Application of Bacillus species under Cr stress increased sugar and proline contents of leaves of wheat [65]. Similarly, under heavy metal stress, bio-inoculation of Rhizobia leguminosarum, Pseudomonas fluorescens, Luteibacter sp, Sinorhizobium meliloti and Variovorax sp exhibited high sugar and proline in Lathyrus sativus.
5. Conclusions
Soil receiving marble effluent had higher Mn content which is harmful for plants. Contaminated soil harbors Mn tolerant bacteria and among these most important were P. eucrina and B. Safensis. Theses bacteria were tolerant to high concentration of Mn found in naturally contaminated soil having marble effluents as well as induced form i.e MnSO4. P. eucrina and B. safensis have a variety of plant growth promoting qualities including phosphate solubilization and IAA and organic acid production. Bio-inoculation of PGPR alone and in consortium effectively reclaimed Mn polluted soil by accumulating high concentrations of Mn in plant parts. Application of these PGPR and their inoculation with other PGPR may be helpful for reclamation of Mn polluted soil and recovery of soil health for agriculture practices. Since the experiment was conducted in a greenhouse, it is recommended that research should be extended to field condition also for attaining better insight of these bacteria.