Chen, Fang, Zhang, Zhang, Wei, and Ping: Directional production of even carbon volatile fatty acids from peanut meal: effects of initial pH and hydraulic residence time
Research
Environmental Engineering Research 2022; 27(4): 210190.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
In this study, peanut meal was used as the substrate for the continuous production of even carbon volatile fatty acids (ECFA) through anaerobic fermentation. First, a continuous stirred reactor (CSTR) was started by adjusting the initial pH to acclimate the related bacteria, and then the hydraulic residence time (HRT) was optimized. The effects of different initial pH and HRT on the potential of ECFA production by peanut meal fermentation, the transformation and utilization of main organic compounds, and the composition and replacement of microbial community structure behind it were investigated. The results showed that ECFA was the dominant factor in the acid production of peanut meal fermentation, and alkaline pH and HRT = 6 d were the more favorable operating conditions for the hydrolysis and acidification of peanut meal. The average concentration of total volatile fatty acids (TVFAs) obtained was 15,249.50 mg/L, and the content of ECFA varied from 88.51% to 96.27%, with an average ratio of 92.55%. This operation condition drove the formation of microbial community structure, mainly including Prepotella, Tepididimicrobium, Anaerosalibacter, and Igatzschineria, which were favorable for ECFA production, reflecting the potential of using peanut meal for targeted production of ECFA.
Peanut is one of the five major oil crops globally, and it is grown all over the world. According to statistics, China is the country with the largest peanut production globally, with an annual total peanut production of about 133,400 tons, accounting for more than 40% of the world and showing an increasing trend year by year [1]. About 60% of peanuts are used to press for oil production, which will produce a large number of peanut meal by-products during processing, with an annual output of nearly 8 million tons [2]. Peanut meal is rich in organic matter, including 48.68% protein, 32.50% polysaccharide, and 0.8% lipid [3]. It has strong digestion and metabolism ability and is often used in the food processing and feed industry [4]. However, due to the pollution of aflatoxin and the loss of nutrients in pressing [5], peanut meal has low added value and utilization rate, so it is often discarded, resulting in a serious waste of resources. Considering the peanut meal for anaerobic fermentation to recover volatile fatty acids (VFAs) is an emerging economically feasible way. VFAs can be used as an external carbon source for nitrogen and phosphorus removal [6] and as feedstock for some high value-added products, such as antimicrobials, corrosion inhibitors, biodiesel, lipids, bioplastics, and degradable plastic polyhydroxyalkanoates (PHAs) [7]. PHAs are biodegradable and compatible and can be fully decomposed and utilized by microorganisms, a broad application prospect [8]. However, the high cost of synthesis of PHAs greatly limits the large-scale production of PHAs (where the cost of carbon source is over 70%). VFAs produced by anaerobic fermentation of peanut meal can be used as a carbon source to synthesize PHAs, effectively reducing the cost of carbon source [9]. Among them, the proportion of even carbon volatile fatty acids (ECFA) is particularly important. Increasing the proportion of ECFA in VFAs can increase the proportion of poly-β-hydroxybutyric acid (PHB) in the PHAs monomer, thus improving the water resistance, thermoplasticity, and biodegradability of the end product [10]. Therefore, in anaerobic fermentation recycling VFAs, targeted to improve the ECFA proportion is of great significance.
pH has a significant effect on the hydrolysis and acidification process of anaerobic fermentation and is one of the decisive factors in the proportion of fermentation end products, especially in terms of acetic acid and butyric acid [11]. It is reported that the pH value of 5.0~11 is beneficial to the production of VFAs. The acetic acid content is usually higher at alkaline pH, while butyric acid is mainly produced at neutral and weakly alkaline pH [7, 12]. In addition to pH, hydraulic retention time (HRT) is another critical parameter that affects fermentation acid production. It determines the organic loading rate and biomass retention and significantly affects anaerobic microorganisms’ metabolic rate [13]. Various studies have pointed out that lower HRT (5–10 d) can prevent methane production growth with a longer generation time [7], improve the hydrolysis capacity of fermentation, and promote the accumulation of ECFA, which requires further optimization. However, there is still a lack of relevant studies in the literature on using different HRTs at alkaline pH on the targeted production of ECFA. In addition, immobilization of microorganisms in the reactor to prolong their residence time, domestication and cultivation of suitable and stable microbial communities are important measures to achieve the accumulation of ECFA. Therefore, continuous flow of production and domestication need to be considered. Peanut meal has strong sedimentation and is easy to be trapped in the reactor, but the mass transfer performance between peanut meal and microorganisms is poor. Continuous stirred reactor (CSTR) of the microbial community has a flocculent structure, increasing the contact area of peanut meal and microorganisms. The operation mode of constant feeding and continuous stirring can improve the mass transfer efficiency of organic matter and realize the continuous degradation and transformation of peanut meal [14, 15].
There are few studies on the production of VFAs by using peanut meal as substrate in CSTR. Especially in regulating the single acid spectrum of VFAs, studies on the acid-producing capacity and microbial community characteristics of specific substrates are not mature. The purpose of this study was to explore the effects of different initial pH and HRT on the directed production of ECFA by anaerobic fermentation of peanut meal in a CSTR and to optimize the operating conditions. In addition, the potential mechanism of ECFA in peanut meal production was discussed from the perspective of organic matter transformation and microbial community. This work may provide valuable theoretical and applied parameter guidance for the directional output of VFAs by solid waste anaerobic fermentation.
2. Materials and Methods
2.1. Raw Materials
The peanut meal comes from an oil mill in Guangzhou. Expose the peanut meal to the sun for 48 h to sterilize and remove the residual moisture in the material, then put it in a food mixer for crushing and sieving (0.4 mm aperture) to obtain uniform powder particles, and store in a dry place for later use. The essential characteristics of peanut meal are (all data are based on wet weight): TS = 99.2%, VS/TS = 91.76%, TOC = 242.71 mg/g, TN = 90 mg/g, C/N = 6.4/1.
To make the organic matter in peanut meal hydrolyze more efficiently and facilitate the production of VFAs in the acid production stage, the pretreated peanut meal supernatant was used as the influent of the reactor in the continuous flow stage. In the previous research of this experiment, to screen out the appropriate pretreatment methods, we used ultrasonic, strong alkali, strong acid, high-temperature strong alkali, and high-temperature strong acid to carry out the pretreatment test of peanut meal. It was found that under high-temperature strong alkali, the hydrolysis solubilization effect of organic matter in peanut meal was the best. The treatment time was short, and the hydrolysis efficiency was high (the data was not given). Therefore, the peanut meal was pretreated with high temperatures and strong alkali. The operation was as follows: 120 g of powdered peanut meal was weighed and placed in a glass container with a sufficient volume of 2 L, then 1.88 L water was added, and the TS concentration was adjusted to 6%. Then, the pH was adjusted to 12 with 3 mol/L NaOH, put into a shaker with a water bath temperature of 55°C, oscillated for 12 h, stood for 12 h, and then the supernatant was taken out for use. The pH of the supernatant was adjusted to 9.0 (obtained from the comparison of two different pH values in the start-up stage), and the chemical oxygen demand (SCOD) was 41,000 mg/L, which was used as continuous inflow water for CSTR.
2.2. Experimental Equipment
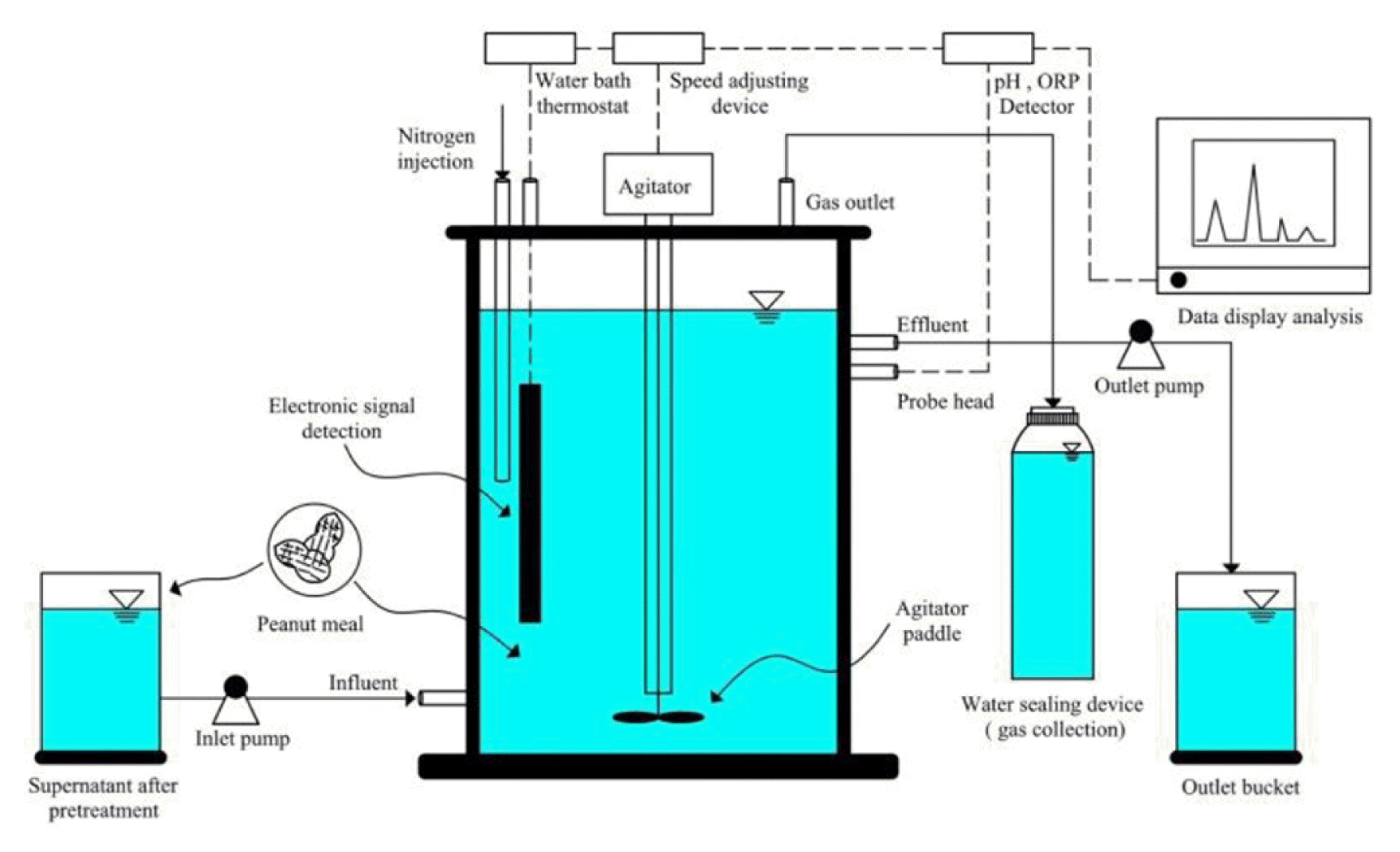
The continuous flow device used in this experiment is a set of CSTR (Fig. 1). The primary material is glass, which has an integrated structure of the reaction zone and precipitation zone, with a total volume of 10 L and a practical volume of 8.5 L. There is a stirring device (rotating speed 40 rpm, periodic 10 min/h) to promote the complete contact and mixing of substrate and water, improve the mass transfer efficiency, and ensure the reaction system’s anaerobic environment through shaft and water seal. The water bath’s constant temperature is adopted, and the reaction temperature is monitored and adjusted at 35 ± 1°C in real-time through the continuous temperature controller. The acid-base changes in the reaction system are recorded by pH and ORP online.
2.3. Experimental Design
2.3.1. Start-up stage
In this experiment, seeding was not carried out. In the start-up stage, the number of hydrolyzed acid-producing microorganisms was increased through the self-fermentation of peanut meal to provide a microbial basis for the subsequent continuous abortive acid. The start-up stage does not set the inlet and outlet water, composed of two parts: the phase without control of pH (start-up stage I, denoting Q1) and the phase with alkaline pH (start-up stage II, denoting Q2), which are respectively run for 11 d. The 510 g peanut meal was put into CSTR at one time, 7.99 L water was added, TS = 6% was adjusted, and the setting temperature was T = 35 ± 1°C. Q1 ferments naturally under this condition. At the end of Q1, the pH value in the reactor was adjusted to 9.0 ± 0.1 with 3 mol/L NaOH, and then Q2 was entered. The reactor was aerated continuously with nitrogen for 15 min before Q1 and Q2 to keep the reactor in an anaerobic state.
2.3.2. Continuous flow stage
At the end of the start-up stage, HRT is gradually regulated in stages, keeping other conditions unchanged. The pretreated peanut meal supernatant is continuously fed into water. To obtain as much ECFA as possible, maintain a better buffer system, only adjust the incoming water’s pH, and do not interfere with the running process’s internal pH. HRT regulation includes three stages. HRT regulation consists of 3 stages: HRT = 8 d (recorded as H8), HRT = 6 d (recorded as H6), and HRT = 4 d (recorded as H4).
In the fermentation process, when the system runs relatively stable during each stage of operation, it enters the next level, during which other working condition parameters remain the same. Each indicator’s changes in the two-stage reactor are recorded daily. A steady-state is considered when the concentration of TVFAs changes by no more than 10%.
2.4. Analytical Method
According to the “Water and Wastewater Monitoring and Analysis method (Fourth Edition)” issued by the National Environmental Protection Agency, various routine analyses are carried out. The analysis items include total organic carbon (TOC), SCOD, ammonia nitrogen (NH4+ – N), total solid (TS), volatile solid (VS) [16]. Total nitrogen (TN) was tested using an elemental analyzer (model EA2400II) manufactured by Perkin Elmer USA. Soluble protein was determined by the Bradford method [17], in which the total protein included granule protein and soluble protein. The concentration of total protein was estimated by multiplying the difference between TN and NH4+ – N concentration by 6.25 [18]. Due to laboratory conditions limitation, only dissolved polysaccharides were determined by the phenol sulfuric acid method [19]. The concentration and composition of VFAs and ECFA were determined by gas chromatography [20]. The VFAs discussed in the article include acetic acid, propionic acid, n-butyric acid, isobutyric acid, n-valeric acid, isovaleric acid, and ECFA contains acetic acid, n-butyric acid, and isobutyric acid. Before analysis, add formic acid to acidify the sample to pH = 3 to reduce volatile acid dissociation’s influence on the results [21]. The gas chromatograph parameters are as follows: the detector is a flame ionization detector (FID), and the chromatographic column is a CNW-WAX capillary column. The carrier pressure is 0.50 MPa for nitrogen, 0.40 MPa for air and 0.20 MPa for hydrogen. The temperature of the oven, sampler, and FID detector are 80, 220, and 250°C, respectively. The oven temperature program is to hold at 80°C for 2 min initially, then at 20°C/min to 110°C for 1 min, and then at 10°C/min to 180°C for 1 min, and the acquisition time is 12.5 min.
2.5. Model Fitting and Statistical Analysis
The establishment of mathematical models can show the biochemical system’s reaction process and provide valuable guidance for optimizing experiments. The concentration in the following formula is expressed as C (mg/L), and the time is shown as t (d).
The degradation rate of organic matter can be calculated according to Eq. (1), where C0 represents the influent concentration and Ct represents the concentration on a certain day(d) in the fermentation process [22].
(1)
Refer to the Anaerobic Digestion Process Model No. 1 (ADMI) of the International Water Quality Association, and use the first-order kinetic model to fit the hydrolysis process to analyze the effect of alkaline pH on the hydrolysis of peanut meal [23], see Eq. (2). vp is the protein hydrolysis rate(mg/(L·d), Cp is the protein concentration on a certain day (d) during fermentation, kh,p is the protein hydrolysis rate constant (d−1).
(2)
The simplified formula is expressed as Eq. (3), where B is the undetermined coefficient.
(3)
The Logistic equation can be used to describe the acidification process, and its expression is as Eq. (4):
(4)
Ci indicates the concentration of VFAs in the fermentation broth at t time, Ci,max is the maximum concentration of VFAs in the fermentation broth, is the maximum specific acid production rate (d−1) “i” can represent TVFAs or ECFA. For further simplified Logistic equation expression, see Eq. (5).
(5)
Origin is used for linear and nonlinear curve fitting, and the Levenberg-Marquardt optimization algorithm is used for iteration. Pearson’s correlation coefficient method is used for correlation analysis to understand the influence of different working conditions on acid production efficiency (Table S2). The coefficient is represented by r (−1 ≤ r ≤ 1), see Eq. (6):
(6)
Among them, , Ȳ and δX are the standard score, the sample mean, and sample standard deviation of Xi, respectively. Use IBM SPSS Statistics 22 software to perform Pearson analysis on the data and use a one-tailed test for significance.
2.6. Microbial Community Analysis
The microbial community analysis test was commissioned by Shanghai Majorbio Bio-pharm Technology Co., Ltd. Under the SmallRNA sequencing analysis platform, DNA extraction and gene amplification were performed on samples under different control conditions, and then 454 pyrosequencing sequencing was performed. Analyze the composition and content of microorganisms in the ECFA production system of CSTR anaerobic fermentation during the start-up and HRT regulation phases. Set up two analysis groups, group 1 is Q1, Q2, H8; group 2 is H6 and H4. Analyze samples in the period of maximum acid production, with two parallel examples in each group. Observe the species diversity index’s changes, structural succession, and microbial community composition at the phylum and genus levels.
3. Results and Discussion
3.1. Effects of Initial pH and HRT on TVFAs and ECFA Accumulation
Under the conditions of natural fermentation and initial pH = 9.0, the CSTR was started to run stably, and the two stages ran for 11 d, respectively. For the changes in TVFAs concentration and components, see Fig. 2(a). It can be seen from the diagram that under steady-state conditions, the average TVFAs level in Q1 and Q2 reactors is 5,724.59 mg/L and 13,615.35 mg/L, respectively. The value of Q2 is 2.4 times higher than that of Q1, indicating that initial alkaline pH can promote the accumulation of VFAs. It is because the collection of VFAs during natural fermentation will lead to systematic acidification. VFAs at low pH exist in a non-dissociated form, which can freely pass through the cell membrane to inhibit the growth of microorganisms [24] and may lead to the denaturation of related enzymes in the system, resulting in a low level of TVFAs production. In contrast, the alkaline environment has a buffer effect on the acidification system, which is conducive to the forward acidification reaction and relieves the inhibition of VFAs accumulation on continuous acid production. Simultaneously, alkaline pH can inhibit methanogens’ activity and reduce the amount of VFAs consumed [25].
The fitting results of the Logistic equation can be found in Table 1. The data shows that the degree of fit in the whole process is low (), but the model in the second half of the reaction has a higher degree of fit (R2 > 0.8). Microorganisms need to have an adaptation time to environmental changes. After the system gradually stabilizes, it begins to show characteristics that conform to the Logistic equation. When the reaction becomes stable, the continuous acid production of peanut meal driven by alkali can refer to the Logistic equation, which has guiding significance for quantifying the peanut meal’s continuous acid production. Interestingly, although both Cmax and acid production have increased significantly; the acid production rate μ has decreased. There is a similar finding that an increase in pH may affect acidification [26]. Besides, the impact of a single model parameter on the logistic fitting cannot be ignored. Therefore, optimizing the model and modifying the relevant fermentation kinetic parameters will explain the dynamic peanut meal anaerobic fermentation of acid production.
During HRT regulation, the three stages ran for 16 d, 14 d, and 12 d. The yield and composition of TVFAs in the reactor were shown in Fig. 2(b). H6 is the most suitable condition for acid production. The average concentration of TVFAs at stable conditions was 15,249.50 mg/L, which was 1.08 and 1.38 times higher than H8 and H4. Suitable HRT conditions allow sufficient time for microorganisms to reproduce and produce acid while avoiding excessive retention of methanogenic bacteria, makes the reactor stay in the acid production section [11].
pH can primarily affect the yield and composition of ECFA (Fig. 2(a)). More acetic acid and butyric acid were observed in the start-up stage, indicating that ECFA dominated the acid production of peanut meal. Under Q1 and Q2, the average proportion of ECFA in TVFAs reached 87.15% and 93.50%, respectively. The alkaline environment not only increased the concentration of VFAs but also increased the percentage of ECFA. With the increase of pH, the proportion of acetic acid increased from 21.2% to 66.0%, and n-butyric acid decreased from 61.3% to 24.4%. The change of isobutyric acid was not visible. In the stage of acid production regulated by HRT (Fig. 2(b)), ECFA is still the leading product; H6 has the highest proportion of ECFA in TVFAs (92.55%), followed by H8 (86.83%) and H4 (87.04%). The effect of HRT change on the proportion between acetic acid and n-butyric acid was less than that in the start-up stage. When H8, H6, and H4 were stable, the proportion of acetic acid was 51.04%, 68.15%, and 54.53%, respectively, and the proportion of n-butyric acid was 32.78%, 22.97%, and 86.75%, respectively.
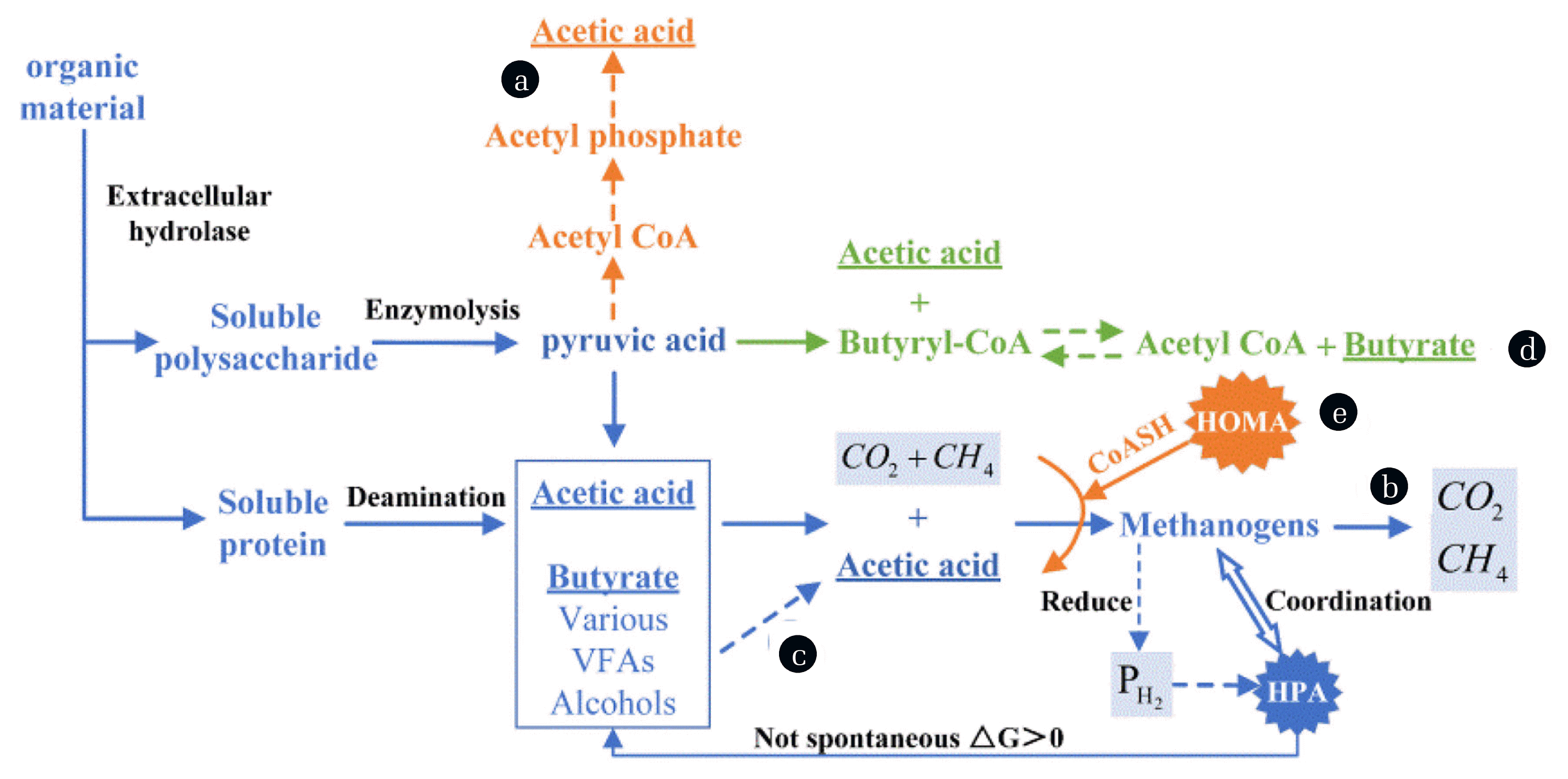
The above results show that whether changing the initial pH or HRT, peanut meal continuous anaerobic fermentation acid production is dominated by ECFA, mainly acetic acid and n-butyric acid. Also, acetic acid dominance in the reaction process may be more determined by the initial alkaline pH. According to literature reports, 65% of methane production comes from butyric acid and acetic acid in VFAs [27]. The loss of methane bacteria with long generations in shorter HRT [28], coupled with the inhibition of methanogenic bacteria activity by alkaline pH [25], makes ECFA have many production opportunities. Acetic acid and butyric acid are the most common ECFA products. Of course, there are more ways to produce acetic acid in most cases, while butyric acid is relatively simple. The typical production routes of VFAs and ECFA are summarized in Fig. 3 [11, 29, 30]. There is a competitive relationship between acetic acid and butyric acid in the production of substrates, which is an important reason why the two acids increase on one side and decrease. Concerning the change in the proportion of acetic acid and butyric acid, the reasons may be attributed to the following:
The formation pathway of acetic acid has no reduction ability, and there are too many acidic end products. In contrast, the formation of butyric acid depends on acetic acid to form acetyl CoA, reducing the acidic terminals. Under the low pH after VFAs accumulation, acid-producing bacteria’s feedback mechanism will increase the conversion rate of butyric acid to maintain the system’s balance, so the acid production of Q1 is dominated by butyric acid and supplemented by acetic acid.
The inhibition of methanogenic bacteria in the alkaline environment made the reaction stay in the non-methanogenic stage, the partial pressure of hydrogen (PH2) in the system increased, the homogeneous acetic acid production process was promoted, and the transformation of butyric acid to acetic acid occurred.
The homogeneous acetic acid production process and some methanogenic bacteria will consume H2. When the PH2 in the system decreases to a certain extent, the hydrogenesis and acetogenesis process will occur, and the transition from butyric acid to acetic acid will also appear.
3.2. Contribution of Organic Matter in Peanut Meal to ECFA Accumulation under Different Initial pH
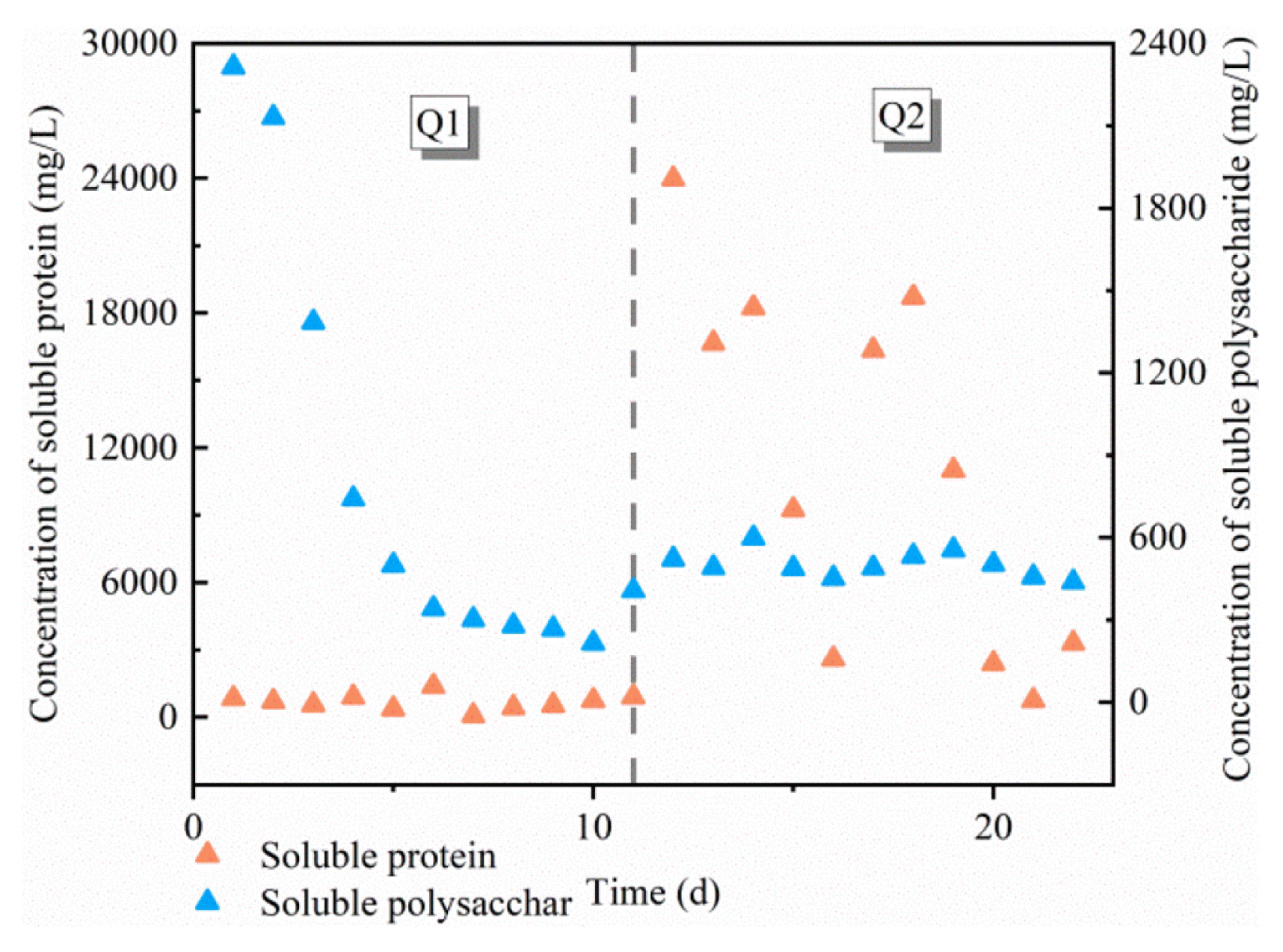
Hydrolysis is generally considered to be the rate-limiting step of anaerobic fermentation. Proteins, polysaccharides are used as the primary nutrient substrates for acid production [31]. This study explored the contribution of organic matter in peanut meal to ECFA accumulation through protein and soluble polysaccharides changes in the start-up stage. The results can be found in Fig. 4.
The polysaccharide structure is more straightforward than protein, and it is easier to be hydrolyzed under natural fermentation. At the beginning of the start-up stage, the soluble polysaccharide concentration is 2.8 times that of soluble protein. As a result, the reaction was more inclined to choose soluble polysaccharides as fermentation substrates, and the concentration of soluble polysaccharides dropped by 78.4% in just five days. Combined with Fig. 3, when pH is not adjusted, the dominant butyric acid is mainly derived from polysaccharide fermentation. The result is similar to the hydrolysis law observed in [32].
Q2 is a better condition for acid production. When stable, the average concentrations of soluble protein and soluble polysaccharides are 2,156.60 mg/L and 466.26 mg/L, almost 2.9 times and 1.6 times that of Q1. The practice of Q2 is similar to alkaline pretreatment, which promotes the dissolution of organic clusters and molecules [33], which facilitates hydrolysis progress. However, the growth rate of the concentration of soluble polysaccharides began to slow down in Q2, and hydrolysis gradually became difficult, which may directly lead to the reduction of butyric acid’s production advantage. On the contrary, the dissolution of protein began to become active. The output and proportion of ECFA increased to a certain extent, becoming the main nutritional contributor to ECFA production. Subsequently, the soluble protein is hydrolyzed into amino acids, and ECFA is accumulated under ammoniation [30]. Correlation analysis showed a significant or extremely significant negative correlation between the concentration of soluble polysaccharides and TVFAs and ECFA under the two initial pH conditions (Q1, P < 0.01; Q2, P < 0.05). Solubility protein concentration showed an extremely significant negative correlation with ECFA concentration only at Q2 (Q1, no correlation; Q2, P < 0.01). Although polysaccharides’ contribution to acid production was slightly reduced, alkaline pH significantly increased protein availability and gradually became the main nutrient contributor, ultimately increasing the total yield of ECFA. The change of the dominant substrate will lead to changes in the composition of the produced acid, which is a fundamental reason for the changes in the proportion of various types of ECFA.
The hydrolysis kinetic model was used to fit the total protein better to understand the hydrolysis process of protein in peanut meal. The relevant results can be found in Table 2. The fitting results show that from Q1 to Q2, kh,p increases from 0.00377 mg/(L·d) to 0.03602 mg/(L·d), which is a 10-fold increase. The initial alkaline pH accelerates the process of protein hydrolysis, establishes favorable conditions for subsequent acid production, and promotes the production of ECFA. Similar to the fitting result of the acid production model, the hydrolysis of protein meets the first-order reaction kinetic equation (R2 > 0.9) after the reaction gradually stabilizes [9]. In addition to the adaptation period required for the response, the deviation of maybe related to the neglect of polysaccharides and lipids in modeling. Other factors, such as particle shape, particle size distribution, and specific surface area, strongly influence hydrolysis processes [18, 34, 35].
3.3. The Utilization of Organic Matter in Peanut Meal by Regulating HRT Acid Production under the Start of Alkaline pH
After pretreatment, the hydrolysis of the peanut meal was effectively strengthened. The influent’s initial soluble protein concentration was 596 = 1.74 mg/L, and the soluble glycogen concentration was 5,641.45 mg/L. Fig. 5(a) shows the observed SCOD, TVFAs/SCOD changes with fermentation time under HRT regulation. The changing trend of the ratio indicates that the SCOD conversion rates of the three stages from high to low are H6 (51.43%), H8 (48.67%), H4 (43.92%), respectively. In H6, organic matter’s solubilization is the strongest, and the SCOD content is the largest. When it is stable, the average value is 30,400 mg/L, 10.7% and 17.8% more than the H8 and H4. Thus, improper HRT may inhibit the solubility of organic matter and prevent the conversion of SCOD to VFAS, resulting from the selection of diverse microbial populations involved in acid production by the fermentation system under different HRT [37]. To further understand the potential mechanism, the microbial community under different HRT will be explored in the following paper.
The degradation rate of soluble proteins and polysaccharides in the fermentation system changed, as shown in Fig. 5(b). When the three stages were stable, the degradation rates of soluble proteins were 83.81%, 72.53%, and 62.26%, respectively. After pretreatment, most of the organic matter has been dissolved out. The degradation rate of soluble protein is gradually reduced, indicating that too short HRT will affect protein hydrolysis, delay amino acidification, and inhibit protein utilization. The difference is that the degradation rate of soluble polysaccharides is always maintained above 90%. Compared with H8 and H4, the concentration of soluble polysaccharides at H6 showed a more significant positive correlation with ECFA yield (H8, no correlation; H6, P < 0.01, H4, P < 0.05), indicating that H6 is more conducive to the solubility of polysaccharides. Thus, when HRT changes, the effect of polysaccharides on the production of ECFA mainly occurs in the hydrolysis stage. However, the impact of HRT on protein utilization primarily happened in the acidification stage because the decrease of soluble protein was negatively correlated with the concentration of TVFAs (H6, P < 0.05; H8 and H4, no correlation). In conclusion, H6 is more conducive to the hydrolysis of polysaccharide and the acidification of protein, which is more conducive to producing ECFA from peanut meal.
3.4. Characteristics of the Microbial Community during Operation
Microbial analysis revealed the effect of different initial pH and HRT on the microbial community in acid production by anaerobic fermentation of peanut meal. Alpha diversity analysis (see Table S1) showed that the microbial community richness in the system was high at the start-up stage, and alkaline pH had little effect on microbial richness (Chao index: Q1,148.6454; Q2. 145.9372). When CSTR enters the continuous flow stage (i.e., H8), the Chao index increases to 155.7502, which may be caused by the enrichment of some microorganisms in the supernatant of peanut meal after pretreatment. However, community abundance decreased with the shortening of HRT due to the increase of water discharge and microbial loss. On the other hand, Shannon and Simpson indices showed an upward trend from Q1 to H8, and a downward trend from H8 to H4, indicating the changing movement of microbial community diversity in CSTR. The results showed that alkaline pH was beneficial to microbial community stability, and appropriate HRT could increase microbial diversity and create conditions for acid-producing fermentation.
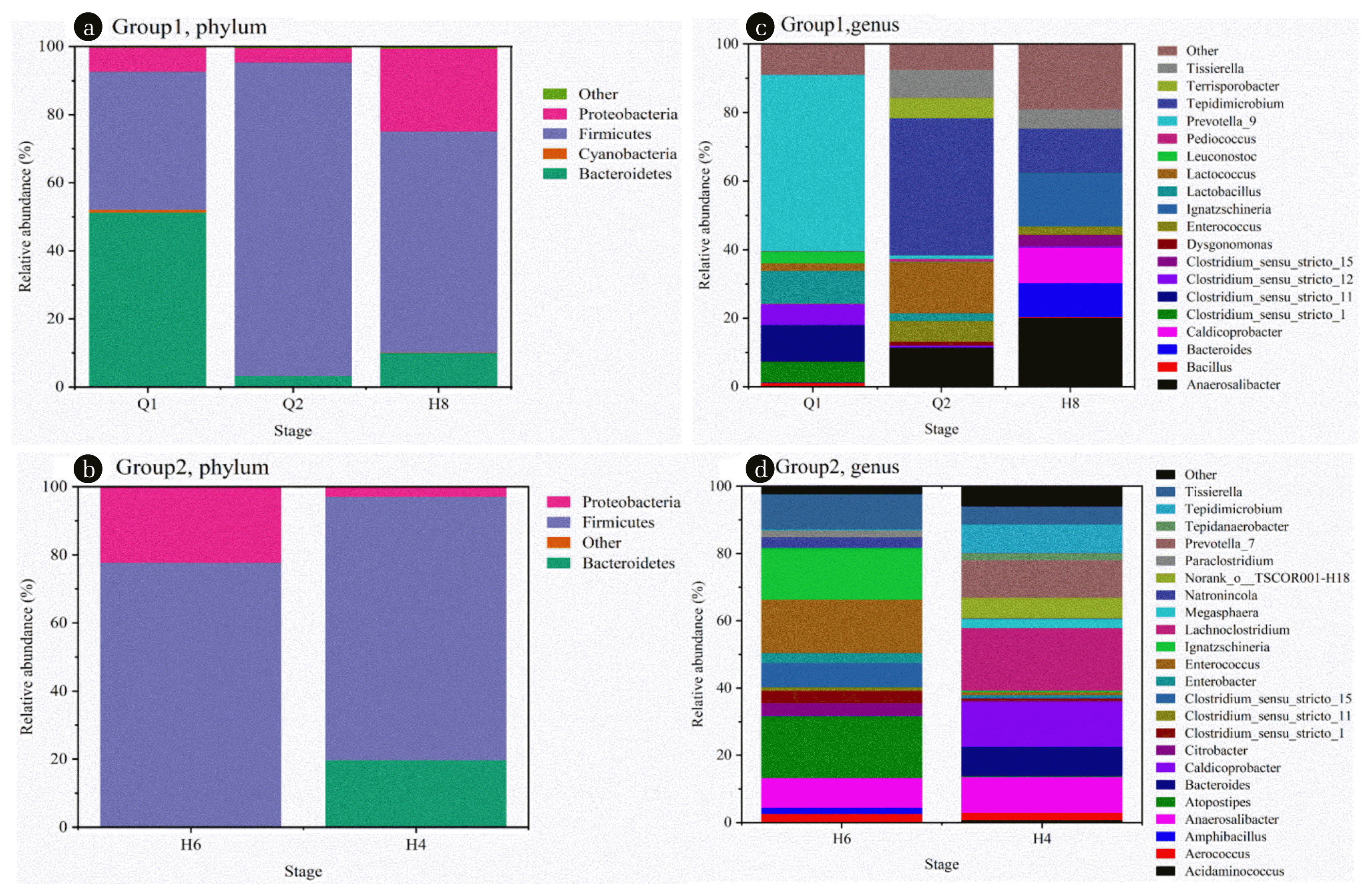
In addition, the results showed the uneven distribution of related fermentation bacteria at the phylum level (see Fig. 6(a), 6(b)). Bacteroidetes and Firmicutes were the two main species in the start-up stage, followed by Proteobacteria. Interestingly, during the transition from Q1 to Q2, Bacteroidetes significantly decreased (51.41% vs. 3.34%) while Firmicutes enriched (40.47% vs. 92.03%) in the community. It is reported that Firmicutes and Bacteroidetes are the most common hydrolytic bacteria in various anaerobic digestion reactors, which are closely related to the hydrolysis and acidification of organic matter [37, 38]. In particular, Firmicutes can metabolize various organics, such as proteins, polysaccharides, lipids, and cellulose [39]. Alkali fermentation increased the content of Firmicutes and the yield of ECFA in the system, suggesting that Firmicutes may be a key player in the acidification process of peanut meal. Bacteroidetes showed a slight disadvantage under alkaline conditions, while Proteobacteria showed a relatively stable abundance under two different pH conditions. Prevotella belonging to Bacteroidetes and Tepidimicrobium belonging to Firmicutes are the dominant bacteria genus in Q1 and Q2, accounting for 51.27% and 40.00%, respectively (Fig. 6(c)). Prevotella can be fermented with polysaccharides to produce VFAs with acetic acid as the main product [40]; Tepidimicrobium is a special genus of bacteria with the ability to decompose proteins, and its end products are still dominated by acetic acid and butyric acid [41]. Their high abundance explains why ECFA is the main product of the anaerobic fermentation of peanut meal. In addition, the replacement of the two dominant bacteria genera also explained the underlying reason for the change in the composition of ECFA in the system by replacing polysaccharide with protein as the main fermentation substrate in Q2.
When CSTR started to enter the continuous flow (H8), Firmicutes decreased to 64.98%, while Bacteroidetes and Proteobacteria increased to different degrees, accounting for 10.21% and 24.29%, respectively. Under the condition of maintaining the inflow and outflow of water, CSTR will affect the microbial community in the system to a certain extent, leading to the instability of the microbial community formation process. Firmicutes, the core flora, still existed in the whole continuous flow stage, suggesting that Firmicutes flora also had important significance for adapting to different HRT in the anaerobic system of peanut meal. However, the content of Bacteroidetes in H6 decreased significantly (< 1%), while Firmicutes and Proteobacteria maintained a relatively high level, with a relative abundance of 77.78% and 22.18%, respectively. With the change of HRT, the unit organic load or water discharge in CSTR changed, and the system showed a difference at the genus level (Fig. 6(d)). It can be seen that the acid-producing bacteria such as Anaerosalibacter, Caldicoprobacter, and Ignatzschineria all increased after entering the continuous flow, accounting for 20.07%, 10.31%, and 15.46%, respectively. During the transition from H8 to H6, the abundance of dominant bacteria Ignatzschineria was relatively stable (15.46% and 15.33%, respectively). With the shortener of HRT, the abundance decreased to 0.41%. Anaerosalibacter and Caldicoprobacter can ferment various sugars and produce organic acids such as lactic acid, acetic acid, and butyric acid. Ignatzschineria produces lactic acid and acetic acid by fermentation of mixed acids [42]. Also, it was observed that the abundance of Clostridium in H6 was 8.25% and 10.98% higher than that in H8 and H4. Clostridium has been identified as a key participant in the acetic acid production process. It is a key hydrolytic bacterium to degrade organic matter into VFAs such as acetic acid [43]. The change of HRT presents different production results of ECFA, which reflects the sum of microbial diversity, dominant flora abundance, and metabolic capacity in the system. These results support the possibility of continuous high concentration ECFA from peanut meal in the anaerobic fermentation system.
4. Conclusions
The results of continuous anaerobic fermentation using peanut meal as substrate were mainly ECFA. The initial pH was alkaline, and HRT was 6 d, which was the most suitable operation condition for the directional production of ECFA from peanut meal. This condition accelerated the hydrolysis and acidification process and promoted the formation of a cooperative microbial community. The presence and change of Prevotella, Tepidimicrobium, Anaerosalibacter, and Ignatzschineria in the acid production system of peanut meal had the most significant effect on ECFA yield.
This work was supported by the Guangzhou University, the National Natural Science Foundation of China (grant no. 2120023), Natural Science Foundation of Guangdong Province (grant no. 2017A030313 273).
Notes
Author Contributions
Y.J.C. (Master student) conducted the experiment and finished this manuscript. Q.F. (Professor) designed the whole experiment and provided laboratory equipment. L.Z. (Master student) participated across the entire investigation. K.Q.Z. (Master student), T.W. (Master student), and Q.Q.P. (Master student) assisted in completing data measurement.
References
1. Fletcher SM, Shi Z. An Overview of World Peanut Markets, in Peanuts. 2016;267–287.
2. Zhan JL, Zhao EL. Research progress on preparation of peptides from peanut meal protein. Agric Technol. 2017;37:7–8.
3. Liu QF, Li ZQ, Jia M, et al. Comprehensive Utilization of Peanut Meal. Food Res Develop. 2017;38:192–195.
4. Zhang Y, Liu J, Lu X, et al. Isolation And Identification Of An Antioxidant Peptide Prepared From Fermented Peanut Meal Using Bacillus Subtilis Fermentation. Int J Food Prop. 2012;17:1237–1253.
5. Mekouar A, Mohamed . 15. Food and Agriculture Organization of the United Nations (FAO). Yearbook of International Environmental Law. 2017;28:506–520.
6. Agler MT, Wrenn BA, Zinder SH, et al. Waste to bioproduct conversion with undefined mixed cultures: the carboxylate platform. Trends Biotechnol. 2010;29:70–78.
7. Fang W, Zhang X, Zhang P, et al. Overview of key operation factors and strategies for improving fermentative volatile fatty acid production and product regulation from sewage sludge. J Environ Sci (China). 2020;87:93–111.
8. Huang L, Chen Z, Xiong D, et al. Oriented acidification of wasted activated sludge (WAS) focused on odd-carbon volatile fatty acid (VFA): Regulation strategy and microbial community dynamics. Water Res. 2018;142:256–266.
9. Yang X. Enhancements of fermentated volatile fatty acids producing by wasted activated sludge and recovery of fermentation liquid. Harbin Institute of Technology; 2012. p. 48–89.
10. Jia Q. Study on synthesis of polyhydroxyfatty acid esters from simulated sludge hydrolysate by complex bacteria. Berijing: Tsinghua University; 2013. p. 87.
11. Ren NQ, Wang AJ, Ma F. Physiological Ecology of Acidogens in Anaerobic Biotreatment Process. Beijing: Science press; 2005.
12. Azman S, Khadem AF, Lier JBV, et al. Presence and Role of Anaerobic Hydrolytic Microbes in Conversion of Lignocellulosic Biomass for Biogas Production. Crit Rev Environ Sci Technol. 2015;45:10.
13. Feng Q, Song Y-C, Kim D-H, et al. Influence of the temperature and hydraulic retention time in bioelectrochemical anaerobic digestion of sewage sludge. Int J Hydrog Energy. 2019;44:2170–2179.
14. Wang WN. Regulation and control of the conversion of CSTR methane fermentation system to acid-producing fermentation system. Harbin: Harbin Institute of Technology; 2008. p. 59–90.
15. Zheng G. Performance and efficient of CSTR and ABR for hydrogen and methane production by organic wastewater fermentation. Harbin: Harbin Institute of Technology; 2010. p. 25–85.
16. Bing T, Passanha P, Kumi P, et al. Recovery and concentration of thermally hydrolyzed waste activated sludge derived volatile fatty acids and nutrients by microfiltration, electrodialysis and struvite precipitation for polyhydroxyalkanoates production. Chem Eng J. 2016;295:11–19.
17. Zhang F, Ouyang P, Zhang X, Chen L. Determination of protein from camel milk by Bradford. Applied Chemical Industry. 2016;45(04)747–750. (in Chinese)
18. Miron Y, Zeeman G, Jules BvL, Gatze L. The role of sludge retention time in the hydrolysis and acidification of lipids, carbohydrates and proteins during digestion of primary sludge in CSTR systems. Water Res. 2000;34:1705–1713.
19. Zheng XW, Huang ZB, Chu DM, et al. Comparison of Phenol-sulfuric Acid and Anthrone-sulfuric Acid Methods for Determination of Polysaccharide in Rubus crataegiflolius Bunge. J Anhui Agric Univ. 2016;044:119–121.
20. Liu ZX. Determination of volatile fatty acids produced by sewage anaerobic digestion by gas chromatography. China Water Supply and Drainage. 2014;30(24)144–146.
21. Chen Q, Liu H, Hu Y, et al. Quick Measurement of Volatile Fatty Acids in Anaerobic Digestion Liquid with Gas Chromatography. China Methane. 2003;21:3–5.
22. Batstone DJ. Mathematical Modelling of Anaerobic Reactors Treating Domestic Wastewater: Rational Criteria for Model Use. Rev Environ Sci Biotechnol. 2006;5:57–71.
23. Li N. The Study of Enhancing SCFAs Production in Waste Actived Sludge Anaerobic Digestion. Nanjing: Nanjing University of Science and Technology; 2015. p. 55–65.
24. Warnecke T, Gill RT. Organic acid toxicity, tolerance, and production in Escherichia coli biorefining applications. Microb Cell Fact. 2005;4:25.
25. Yuan H, Chen Y, Zhang H, et al. Improved Bioproduction of Short-Chain Fatty Acids (SCFAs) from Excess Sludge under Alkaline Conditions. Environ Sci Technol. 2006;40:2025–2029.
26. Ma H, Chen X, Liu H, et al. Improved volatile fatty acids anaerobic production from waste activated sludge by pH regulation: Alkaline or neutral pH? Waste Manag. 2016;48:397–403.
27. Mamimin C, Prasertsan P, Kongjan P, et al. Effects of volatile fatty acids in biohydrogen effluent on biohythane production from palm oil mill effluent under thermophilic condition. Electron J Biotechnol. 2017;29:78–85.
28. Xiong H, Chen J, Wang H, Shi H. Influences of volatile solid concentration, temperature and solid retention time for the hydrolysis of waste activated sludge to recover volatile fatty acids. Bioresour Technol. 2012;119:285–292.
29. Pasteur L. On the viscous fermentation and the butyrous fermentation. Bull Soc Chim Biol (Paris). 1861;284–290.
30. Hu JC. Theory and technology of anaerobic biological treatment of wastewater. Beijing: China Architcture & Building Press; 2002. p. 54–77.
31. Dahiya S, Sarkar O, Swamy YV, et al. Acidogenic fermentation of food waste for volatile fatty acid production with co-generation of biohydrogen. Bioresour Technol. 2015;182:103–113.
32. Wang K, Yin J, Shen D, et al. Anaerobic digestion of food waste for volatile fatty acids (VFAs) production with different types of inoculum: effect of pH. Bioresour Technol. 2014;161:395–401.
33. Lin L, Li X-y. Effects of pH adjustment on the hydrolysis of Al-enhanced primary sedimentation sludge for volatile fatty acid production. Chem Eng J. 2018;346:50–56.
34. Hills DJ, Nakano K. Effects of particle size on anaerobic digestion of tomato solid wastes. Agricultural Wastes. 1984;10:285–295.
35. Hobson PN. A Model of Some Aspects of Microbial Degradation of Particulate Substrates. J Ferment Technol. 1987;65:431–439.
36. Romero Aguilar MA, Fdez-Güelfo LA, Álvarez-Gallego CJ, Romero García LI. Effect of HRT on hydrogen production and organic matter solubilization in acidogenic anaerobic digestion of OFMSW. Chem Eng J (Lausanne). 2013;219:443–449.
37. Lv Z, Wang J, Chen Z, et al. Temperature regulations impose positive influence on the biomethane potential versus digesting modes treating agricultural residues. Bioresour Technol. 2020;301:122747.
38. Lv Z, Wu X, Zhou B, et al. Effect of one step temperature increment from mesophilic to thermophilic anaerobic digestion on the linked pattern between bacterial and methanogenic communities. Bioresour Technol. 2019;292:121968.
39. Liu X, Xu Q, Wang D, et al. Unveiling the mechanisms of how cationic polyacrylamide affects short-chain fatty acids accumulation during long-term anaerobic fermentation of waste activated sludge. Water Res. 2019;155:142–151.
40. Sakamoto M, Benno Y. Reclassification of Bacteroides distasonis, Bacteroides goldsteinii and Bacteroides merdae as Parabacteroides distasonis gen. nov., comb. nov., Parabacteroides goldsteinii comb. nov. and Parabacteroides merdae comb. nov. Int J Syst Evol Microbiol. 2006;56:1599–1605.
41. Niu L, Song L, Liu X, et al. Tepidimicrobium xylanilyticum sp. nov., an anaerobic xylanolytic bacterium, and emended description of the genus Tepidimicrobium. Int J Syst Evol Microbiol. 2009;59:2698–2701.
42. He Q, Li L, Zhai L, et al. Micro-ecological characteristics of dry anaerobic digestion sludge expansion of kitchen waste. Chinese Environ Sci. 2018;38:1010–1017.
43. Lv Z, Chen Z, Chen X, et al. Effects of various feedstocks on isotope fractionation of biogas and microbial community structure during anaerobic digestion. Waste Manag. 2019;84:211–219.
Fig. 1
CSTR device structure diagram.
Fig. 2
Changes in the proportion of ECFA in the TVFAs in the reactor under different initial pH (a) and HRT (b) The main types examined by ECFA are acetic acid, isobutyric acid, and n-butyric acid.
Fig. 3
Describes the typical production pathways of VFAs and ECFA: (a) Hydrogenesis and acetogenesis process (glycolysis); (b) Anaerobic fermentation general acid production process; (c) Other organic matter hydrogenesis and acetogenesis process(-producing acetogens, HPA) (d) Butyric acid production process; (e) Homogeneous acetic acid production process (homo-acetogens, HOMA).
Fig. 4
Changes in the concentration of soluble polysaccharides and soluble proteins during the start-up stage.
Fig. 5
(a) The concentration of SCOD, the value of TVFAs/SCOD, and (b) the degradation rate of the main dissolved organic matter change under the three control stages of HRT.
Fig. 6
The relative abundance of the community at the microbial genus level of the samples of group 1 (a) and group 2 (b); the relative abundance of microorganisms in group 1 (c) and group 2 (d) at the phylum level.