1. Introduction
Microbes can attach to the surface of membranes used in membrane-based water treatment systems and produce polymeric substances that aid in antimicrobial resistance [1]. Extracellular polymeric substances (EPS) are polymeric substances produced by microbes that, when combined with microbial cells, form biofilms. Therefore, biofouling is the development of biofilms in membrane-based water treatment. Biofouling is regarded as the most problematic type of fouling in membrane-based water treatment systems. This is due to the recurrence of biofouling, which adversely affects the life of the membrane, the quality of the water produced, and the efficiency of the system [2–4]. Attempts to control biofouling increase the use of chemicals, which can be harmful if released into the environment indiscriminately [5,6].
Preventing early bacterial adhesion is a promising approach to controlling biofilm formation [7,8]. This could be accomplished by developing antifouling membranes (membranes that prevent the adhesion of microorganisms) [9], antimicrobial membranes (membranes that have the ability to inactivate microbial cells) [10], or dozing the bulk medium with biocides [11–13]. Membrane modification approaches using antimicrobial agents have been used to control biofouling in membrane-based water treatment systems [14–16]. This is done to reduce the toxic chemical waste generated during the water treatment process. The accumulation of organic matter, dead microbial cells, and other substances on the surface of the modified membrane, on the other hand, limits its effectiveness and efficacy. Surface conditioning, for example, is the first step in biofilm development. Surface conditioning is the accumulation of organic materials on the surface of the membrane, creating an environment suitable for microbial attachment. During this process, the conditioning layer can prevent microbes from coming into contact with the antimicrobial agent immobilized on the membrane, thereby inhibiting antimicrobial attacks while also acting as a food source for microbial cells.
Conventional antimicrobial agents such as chlorhexidine, silver salts, quaternary ammonium surfactants, etc. have been used to eradicate biofilms on surfaces. However, antimicrobial peptides (AMPs) have recently received more attention. This is due to their broad antimicrobial spectrum and antimicrobial mode of action, which microbes find difficult to resist. AMP is typically a small cationic structure with a different amino acid composition comprising of distinct hydrophobic and hydrophilic domains. Over the years, AMPs have been immobilized on biomedical surfaces to prevent biofilm colonization [17–19]. A similar approach was used in water treatment applications, where AMPs were attached to the membranes [20,21]. However, when compared to other antimicrobial agents, the challenges of immobilizing AMPs on surfaces include (1) orientation specific activity (AMP can be easily readjusted by the surface energy and its potent orientation distorted), (2) degradation of AMP by environmental enzymes, (3) risk of attachment of AMP to other materials other than the microbial cells due to electrostatic interaction, (4) discovery and synthesis costs, and (5) pH and/or salinity dependent bioactivity [22,23].
Therefore, improving the environmental stability of the AMP (i.e., enzyme, salinity, and pH tolerance) through some modifications and making it soluble in the feed water is a promising method to overcome these shortcomings and achieve higher efficacy. When a stable AMP is added to the feed water, its mobility increases, allowing it to reach the microbial cell wall and penetrate the microbial cell membrane. HHC-36 AMP is a small 9-amino acid with a high charge density. HHC-36 AMP is less expensive than some other AMPs due to its shorter amino acid sequence [24]. HHC-36 has previously been immobilized on biomaterials for biofilm control; however, as far as we know, HHC-36 has never been used for biofilm control in membrane-based water treatments. To improve the stability of HHC-36, the N-terminus was modified with L-proparglycine and tested against Pseudomonas aeruginosa PAO1, an important membrane biofouling bacterium [25–27], both with and without organic matter in the form of a growth medium.
2. Materials and Methods
2.1. Materials
GL Biochem Ltd (China) synthesized the HHC-36 AMP modified with L-propargylglycine at the N-terminus to make it salt and pH tolerant (seq:Pra-Lys-Arg-Trp-Trp-Lys-Trp-Trp-Arg-Arg, Pra-KRWWKWWRR). Phosphate buffered saline (PBS) powder and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich (USA). For the membrane biofouling test, a forward osmosis (FO) membrane (polyamide thin film composite (PA-TFC), CSM FO-8040) from Toray Chemical Korea Inc. (Korea) was used.
2.2. Bacterial Preparation
Single colonies of P. aeruginosa PAO1, Bacillus sp or E. coli were cultured in LB broth for 16 h at 37 °C in a shaking incubator at 150 rpm. The bacterial cultures were then centrifuged for 10 min at 8,000 rpm to wash the bacterial cells. The culture medium was removed and replaced with PBS before centrifuging for 5 min at 8,000 rpm. This last procedure was repeated, and the final concentration of the bacterial cells was adjusted to an optical density of 0.2 at 600 nm wavelength (OD600 0.2) using an Ultrospec 3100 pro UV/Vis spectrophotometer (Amersham Bioscience, USA). The bacterium, P. aeruginosa PAO1 was solely used as a model for biofilm formation on water treatment membranes.
2.3. Effect of Dissolving Solvent on the Antimicrobial Activity of HHC-36 AMP
Organic solvents are commonly used to dissolve AMPs, particularly hydrophobic and neutral AMPs [28]. However, when reporting antimicrobial properties of such AMPs and even some conventional antibiotics, the effects of organic solvents are frequently overlooked. In this study, we look at how dissolving solvents affect the antimicrobial property of HHC-36. The antibacterial property of HHC-36 was tested against Pseudomonas aeruginosa PAO1, Escherichia coli, and Bacillus sp. by dissolving the peptide in (DMSO) (as suggested by the manufacturer), phosphate buffered saline (PBS) and autoclaved deionized (DI) water.
To test the antibacterial activity of HHC-36 AMP, a stock solution was first prepared. 1 mg of HHC-36 AMP was dissolved in 0.5 mL of DMSO, PBS, and autoclaved DI water to make the stock solution. LB broth was used to adjust concentrations to 0 μg/mL, 31.3 μg/mL, 62.5 μg/mL, 125 μg/mL, 250 μg/mL, 500 μg/mL, and 1000 μg/mL using LB broth. 1:1 bacterial cell to LB broth (as prepared above), resulting in final AMP concentrations of 0 μg/mL, 15.7 μg/mL, 31.3 μg/mL, 62.5 μg/mL, 125 μg/mL, 250 μg/mL, and 500 μg/mL, respectively. The cells were then cultured for 24 h at 37 °C and 150 rpm. The optical density of the culture was measured at 600 nm after 24 h using UV/Vis spectrophotometer (Amersham, USA). The baseline was bacterial inoculated LB culture, and the positive control was medium lacking HHC-36 AMP.
2.4. Inhibition of Bacterial Adhesion to FO Membrane by HHC-36
In this study, we investigated the effect of organic matter in the form of bacterial culture medium and without organic matter in the form of PBS on the antimicrobial property of HHC-36. HHC-36 was dissolved in both LB broth and PBS to make a stock concentration of 2 mg/mL in each. Diluting with LB broth or PBS, different concentrations of each condition (0 mg/mL, 0.25 mg/mL, 0.5 mg/mL, and 1 mg/mL) were prepared. 1% and 50% of the prepared bacterial cells were added to organic matter-containing and organic-free samples, respectively. The FO membrane (1 cm × 1 cm) was sterilized by soaking in 70% ethanol, drying it, and then attaching to a 24-well plate (SPL Life Science, South Korea) with the polyamide active side facing up. Following that, 2 mL samples containing bacterial cells, HHC-36 AMP with and without organic matter were added to each well and incubated in a shaking incubator at 37 °C and 150 rpm.
After a 24 h incubation period, attached bacteria on the FO membranes were enumerated using colony forming unit counts, as described in a previous study [29–31]. Briefly, the bacterial suspension was removed from the wells of the cell culture plate and the membranes were rinsed with PBS to remove unattached cells. The FO membranes were detached and placed in 1.5 mL microtubes containing 1 mL PBS. Bacteria that had become adhered to the membrane were detached by vortexing for 2 mins and sonicating for 1 min. The bacterial suspension was serially diluted and cultured on an LB agar plate in an incubator at 37 °C for 24 h. The colonies on the plate were counted after 24 h of culture.
Bacteria attached to the membrane were also analyzed using the Live/Dead Bacterial Viability Kit (Invitrogen Life Technologies, USA). The kit contains a red stain (propidium iodide) and a green stain (SYTO9) which were diluted in filtered autoclaved DI water by adding 3 μL of each stain to 1 mL of filtered autoclave DI water to prepare a working solution. After rinsing the FO membrane with PBS to remove unattached cells, it was stained with 500 μL of the working solution and incubated in the dark at room temperature for 20 mins. Fluorescent pictures of the bacterial cells were taken with a confocal laser scanning microscope (Olympus FV1000, Olympus Inc., USA).
The morphology of the attached bacterial cells on the FO membranes was examined by immobilizing the attached cells with 5% glutaraldehyde (Sigma-Aldrich, USA) and dehydrating with 100% ethanol. Images of the bacterial cells were visualized with a field emission scanning electron microscope (FE-SEM) (Hitachi S-4700, Japan).
2.5. DNA Apoptosis Test
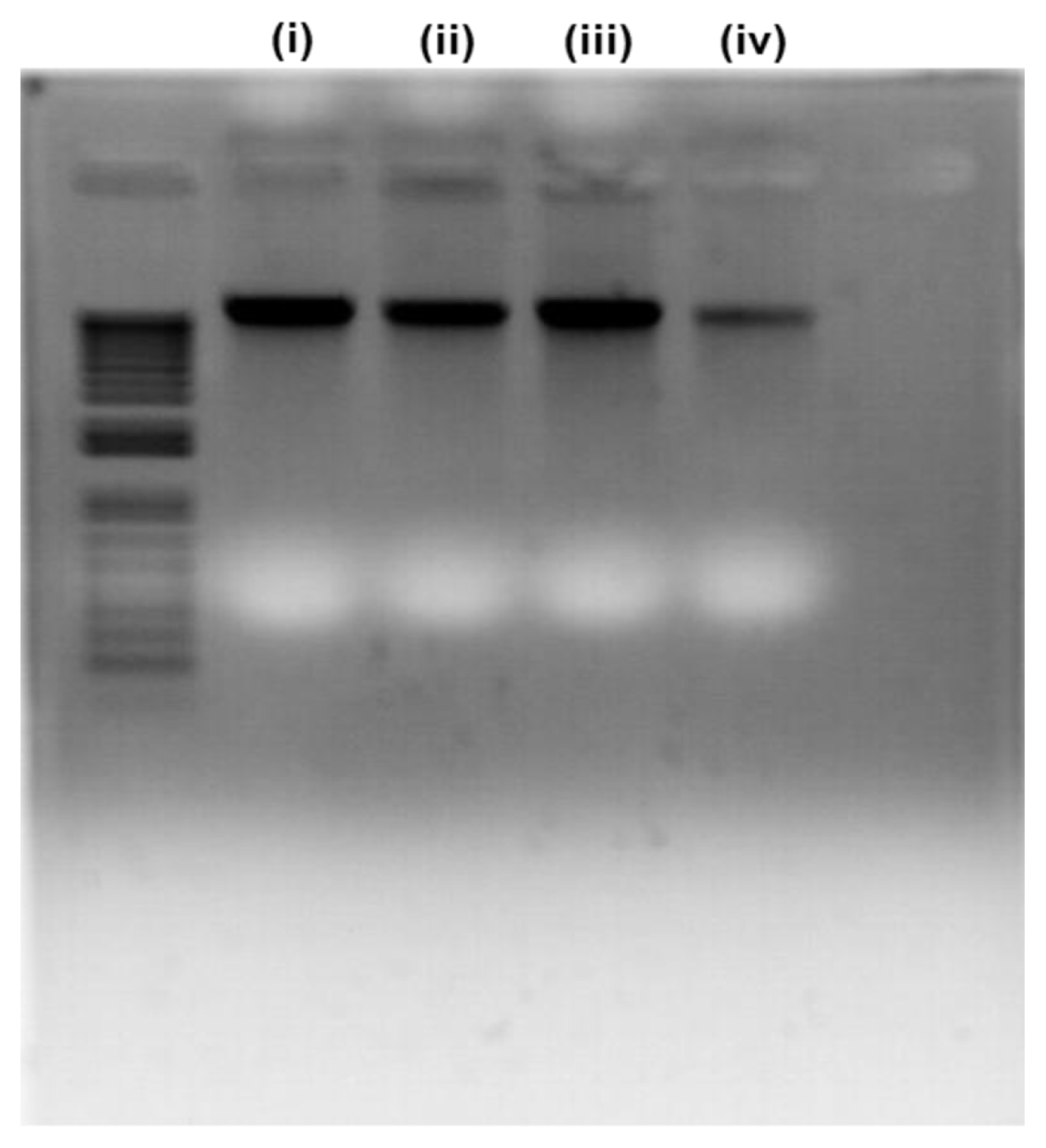
The DNA of the attached bacterial cells (without organics) to the FO membranes was extracted using the AccuPrep® Genomic DNA Extraction Kit (Bioneer Corporation, South Korea) by following the procedure provided by the manufacturer. Bacterial DNA bands were separated on a 2% agarose gel consisting of ethidium bromide. The DNA band was visualized with a UV trans illuminator.
3. Results and Discussion
3.1. Effect of Dissolving Solvents on the Antibacterial Activity of HHC-36
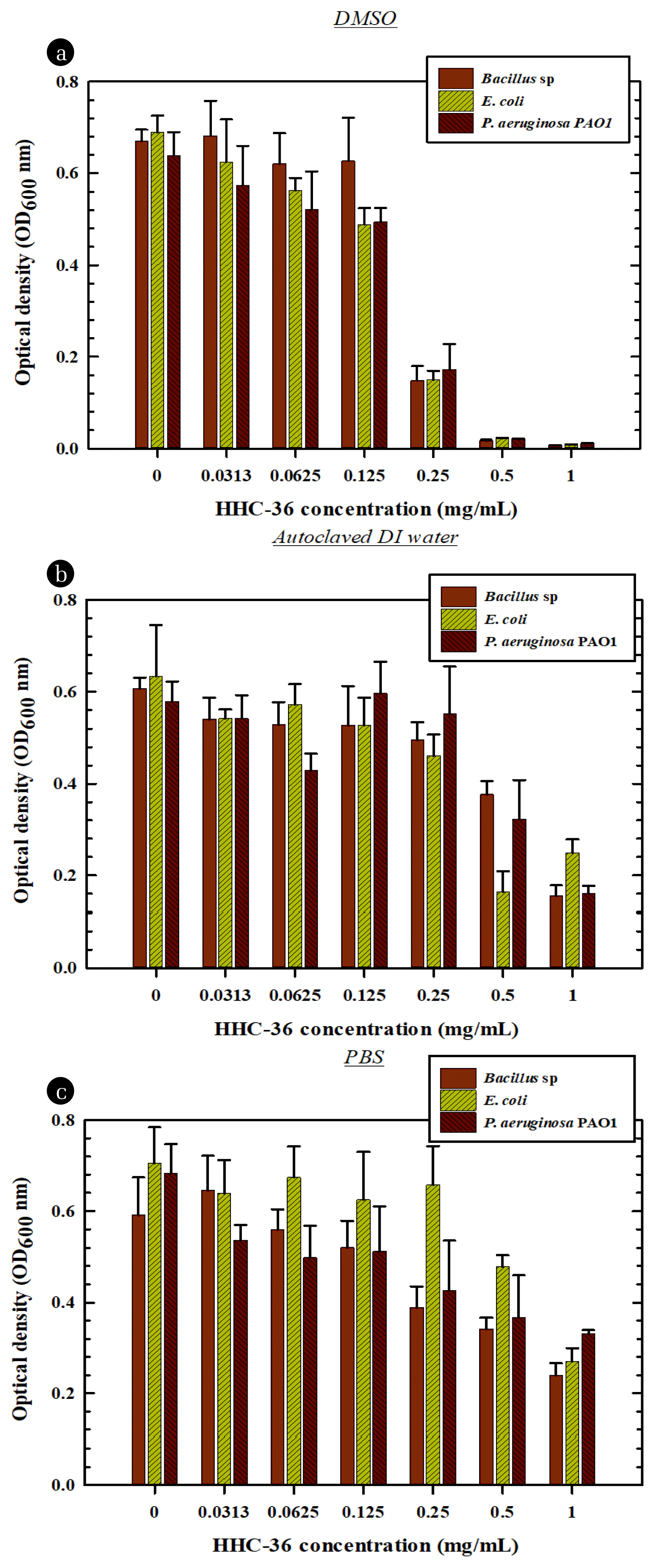
The effect of the solvents used in the stock preparation of the HHC-36 AMP was tested using DMSO, PBS, and autoclaved DI water. Fig. 1 depicts bacterial growth after culturing at various AMP concentrations. The efficacy of the HHC-36 AMP was higher when the peptide was initially dissolved in DMSO. The antibacterial effect increased as AMP concentration increased, with 250 μg/mL and 500 μg/mL being the most effective concentrations. For all bacterial species, the potency of the AMP was reduced when the stock solution was prepared in autoclaved PBS and DI water instead of DMSO.
Several studies have reported the antibacterial activity of HHC-36 [32–34]. The antibacterial mechanism of the antimicrobial HHC-36 is not fully understood. Nonetheless, the mode of antimicrobial action of HHC-36 has been suggested to be a function of its interaction with the bacterial membrane or its ability to penetrate bacterial cells and attack cytoplasmic components [33]. In this study, increasing the concentration of HHC-36 in all solvents suppressed bacterial growth.
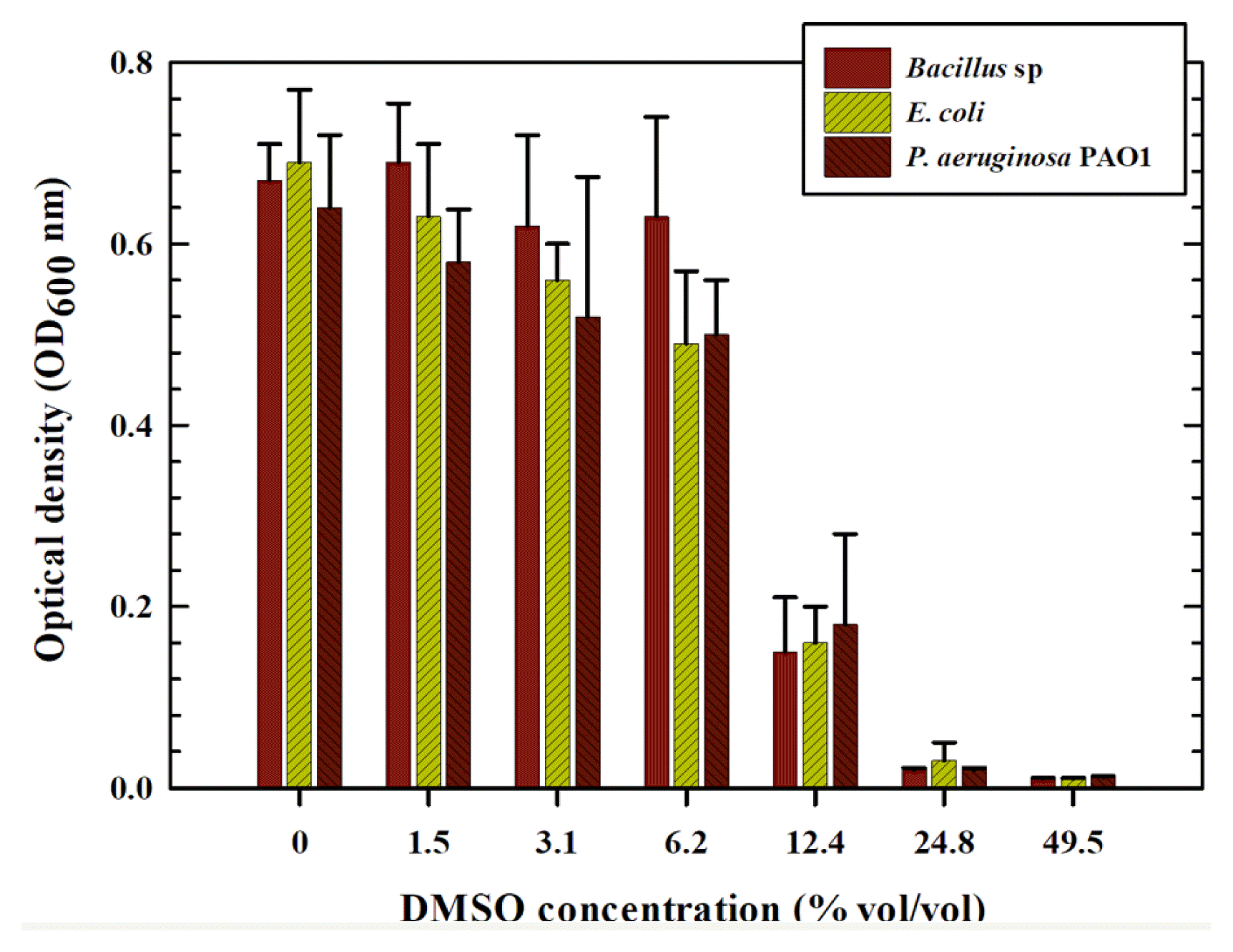
To investigate the potential effects of DMSO on the antibacterial properties of HHC-36 AMP, a serial dilution of the DMSO (without AMP) was prepared and tested against the three bacterial species. Surprisingly, the higher the DMSO concentration, the better the antibacterial properties, as also observed when AMP was dissolved in DMSO (Fig. 2). However, at lower concentrations, the antibacterial activity of the AMP-containing solvent differed slightly. DMSO containing AMP demonstrated greater antibacterial activity than low concentrations of pure DMSO solvent. These results demonstrate the impact of an organic solvent (DMSO) on the antibacterial properties of HHC-36 AMP. This finding is consistent with previous research on the effect of solvents on bacterial growth when determining the minimum inhibitory concentration (MIC) of antimicrobial agents. Wadhwani et al. [35] investigated the susceptibility of various types of bacteria to DMSO and other solvents. All bacterial cells examined in DMSO ≥ 4% in their report, showed a significant reduction in growth. In a related study by Guo et al. [36], the effect of DMSO was tested at non-inhibitory concentrations. The authors found that a non-inhibitory concentration of DMSO (2%, vol/vol) inhibits quorum sensing (QS), thereby attenuating QS-driven virulence factors in P. aeruginosa. They concluded that combining DMSO with antibiotics such as ciprofloxacin, increased P. aeruginosa susceptibility to such antibiotics.
The efficacy of the peptide was lowest when dissolved in PBS. This could be due to the possibility of cationic AMP interference caused by the addition of ions present in the PBS. Ions can bind to AMP, reducing its bioavailability to bacterial cells.
3.2. FO Membrane Biofouling Inhibition
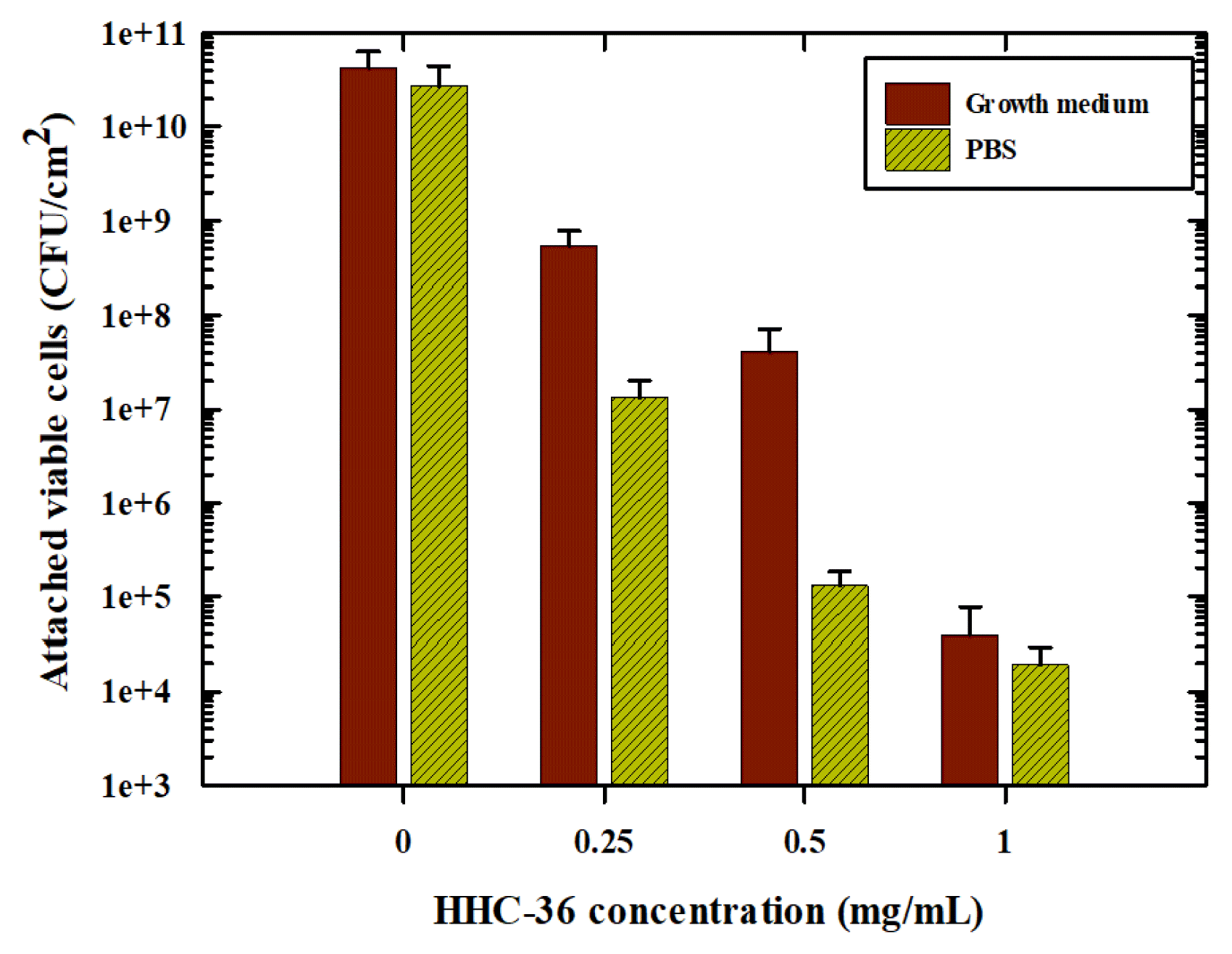
Bacterial growth and adhesion on FO membranes were conducted using P. aeruginosa PAO1. The inhibitory effect of various concentrations of the HHC-36 were tested while autoclaved DI water was used to represent the absence of HHC-36 AMP. There were two conditions tested: inhibition in the presence and absence of a growth medium. Because microorganisms, especially bacteria, do not exist in isolation in the environment, we hypothesized that organic matter could block the reactive sites of HHC-36 AMP, rendering it inactive with bacterial cells, and that the availability of nutrients to bacterial cells could influence microbial susceptibility to HHC-36 AMP. As illustrated in Fig. 3, the efficacy of HHC-36 increased with increasing HHC-36 AMP concentration in the presence or absence of nutrients. This implies that although ionic species may impact the efficacy of HHC-36 AMP as discussed earlier, organic matter in the form of nutrients does not interfere with the activity of HHC-36 AMP. The slight difference in cell numbers observed in the presence and absence of organic matter could be attributed to bacterial growth caused by nutrient availability. Therefore, the proliferation of bacterial cells in the bulk solution indicates that more cells are likely to interact with the membrane surface and eventually adhere.
Fig. 4 shows the live/dead assay for bacterial cells attached to the FO membranes. More dead cells are found on the membranes as the concentration of HHC-36 AMP increased in the presence of a growth medium. Similarly, as HHC-36 AMP concentrations increased, so did the number of dead bacterial cells on the membranes without a growth medium. The results of this assay are consistent with the CFU counts, indicating that as HHC-36 AMP concentration increased in the presence and absence of a growth medium, there were fewer living bacteria adhering to the FO membranes.
FE-SEM images further show the ability of HHC-36 AMP to inhibit bacterial adhesion (Fig. 4 (c) and (d). The bacterial cells incubated with 0 mg/mL HHC-36 AMP show intact membrane structure (Fig. 4 (c–i)). Bacterial cells were attached to the FO membranes in the presence of HHC-36-containing growth medium despite some compromises in cell membrane integrity (Fig. 4 (c, ii–iv). This result is consistent with the work of He et al. [37] involving the immobilization of AMP on the silicon surface. A certain number of bacteria are present in the sample in the absence of a growth medium, and these bacteria can be inactivated by HHC-36 AMP. Some bacterial cells retain the integrity of their membranes, while others were disrupted (Fig. 4 (d, ii–iv)). This result agrees with the CFU and live/dead analyses, which revealed that the bacteria were not completely inactivated. AMPs are thought to inactivate bacteria by penetrating and disrupting cell membranes via a hydrophobic interaction between their cationic nature and the bacterial lipid bilayer [38]. AMPs are drawn to the lipid bilayer by hydrophobic interactions, allowing them to pass through the bacterial membranes and leave pores. Furthermore, the cationic nature of the AMP initiates a counterion exchange via the outer lipopolysaccharide layer of Gram-negative bacteria. This enhances AMP penetration into the bacterial cytoplasm [39,40].
3.3. Bacterial DNA Fragmentation
The potential DNA fragmentation by HHC-36 AMP was investigated on the attached bacterial cells on the FO membranes. Interestingly, the DNA of the attached bacterial cells was intact, with no visible signs of fragmentation (Fig. 5). However, the band thickness of the bacterial cells treated with 1 mg/mL was smaller than that of the other concentrations. Several runs on the agarose gel produced similar results. The reason for the thinner band of cells in contact with 1 mg/mL HHC-36 AMP could not be explained in this study. Notwithstanding, based on our findings, bacterial cell inactivation could not be linked to DNA fragmentation in bacteria.
4. Conclusions
This study first evaluated the effect of initial dissolving solvents such as DMSO, DI water and PBS on the antibacterial activity of HHC-36, which was modified at the N-terminus with L-propargylglycine to allow salt tolerance. At 0.5 mg/mL, the HHC-36 AMP exhibited a significant antibacterial activity regardless of the solvent used to prepare the stock solution. However, preparing the stock in DMSO increased the antibacterial activity of HHC-36 AMP. The use of PBS and DI water reduced the effectiveness of AMP, with PBS having the lowest effectiveness. Some of the most potent antibiotics and antimicrobials are insoluble in water and must be dissolved in organic solvents. However, the potential toxicity of the solvents should be tested to accurately demonstrate the antimicrobial properties of antimicrobial agents requiring the use of solvents. Subtle effects of these solvents, such as interference with genetic expression, need to be considered. This assessment may seem tedious, but it will help in the accurate reporting of antimicrobial agents and reduce the amount of controversy reported in the literature. Treatment with HHC-36 AMP inhibited bacterial adhesion and biofilm formation on the water treatment membrane by disrupting bacterial cell morphology in the presence or absence of nutrients. Nonetheless, the antimicrobial activity of HHC-36 needs to be evaluated in a more complex and diverse environment. More research is needed to determine the impact of HHC-36 AMP on FO membrane performance such as permeate flux, rejection, and reverse solute flux etc. Future studies would also compare the efficacy of HHC-36 AMP in feed bulk solution and immobilized on a membrane to study its biofilm control ability and impact on membrane performance. Overall, this work further strengthens the antimicrobial and potential antiadhesion properties of HHC-36 AMP on the water treatment membranes.