1. Introduction
Methane (CH4), which is mainly produced by methanogens in anaerobic environments, is widely distributed in nature [1, 2] and accounts for almost 20% of global warming every year [3]. It has been reported that the methane produced in wetlands alone is as high as about 1.03 × 108 tons per year [4], but most of which are consumed before being released into atmosphere [5]. On the other hand, wastewater treatment plants (WWTPs), regarded as the emission sources of greenhouse such as carbon dioxide (CO2) and CH4, have also attracted increasing attention [6, 7]. It was reported that about 52,600 tons of CH4 were emitted from the WWTPs of China in 2012 [8]. Meanwhile, a lot of efforts have been made to solve the shortage of organic carbon sources in biological denitrification of wastewater [9]. Recently discovered denitrifying anaerobic methane oxidation (DAMO) not only suggests a link of carbon and nitrogen cycles, but also presents a potential bioprocess for the synchronous removal of nitrite and dissolved methane in anaerobic condition [10, 11]. Raghoebarsing and coworkers discovered that some bacteria could oxidize methane to carbon dioxide (CO2) while reducing NO2− to nitrogen gas (N2) in anaerobic environment, which has been defined as nitrite-dependent anaerobic methane oxidation (n-DAMO) as Eq. (1) [10].
High-throughput sequencing is widely used to analyze the microbial community and understand microbial function and diversity with high accuracy and low cost. It is carried out through sequencing by synthesis and the exclusive reversible termination of chemical reactions which could identify most kinds of microbes in the environment [12]. With the help of high-throughput sequencing, Candidatus Methylomirabilis oxyfera could be first identified from canal sediments [13, 14]. More and more n-DAMO bacteria, such as Candidatus Methylomirabilis sinica, Candidatus Methylomirabilis limnetica and Candidatus Methylomirabilis lanthanidiphila have also been found in the enrichment culture of paddy soil [15], deep stratified lake [16] and ditch sediment [17], respectively. All of the species in Candidatus Methylomirabilis have been affiliated to the phylum of NC10. Radioisotope tracer of 14C revealed that anaerobic methane oxidation was coupled to denitrification by denitrifying methanotrophs as illustrated as Eq. (1) [18]. Candidatus Methylomirabilis has also been detected from various environments such as WWTPs [19], the sediment of lake and river [20], coastal sediment [21], and paddy-peatlands [22]. Obviously, n-DAMO bacteria are widely distributed in nature.
Though any pure cultures of n-DAMO bacteria have not been obtained so far, a few efforts have been contributed to enrich n-DAMO bacteria in wastewater treatment systems for denitrification [23]. With the mixture of lacustrine sediment and activated sludge as inoculum, an enrichment culture of n-DAMO bacteria had been obtained after being enriched for 260 d at 22°C, in which 15% of bacteria were identified as Candidatus Methylomirabilis [24]. Another enrichment culture dominated by Candidatus Methylomirabilis oxyfera was also obtained from anaerobic sludge after acclimatization for about 300 d, which presented a well n-DAMO phenomenon at ambient temperature (20–23°C) [19]. To understand the effect of reactor configuration on n-DAMO process, magnetic stirred gas lift reactor (MSGLR), sequencing batch reactor (SBR), and continuous stirred tank reactor (CSTR) had been evaluated by an enrichment culture of n-DAMO bacteria [25]. A membrane biofilm reactor (MBfR) was also performed for 24 months to demonstrate the feasibility of anammox coupled to n-DAMO for nitrogen removal from wastewater [26]. Inoculated with the sludge collected from a denitrifying bioreactor, n-DAMO activity in a membrane bioreactor could be observed after the acclimatization period as long as 388 d [11]. It has also been reported that via the enhancement of conductive materials, n-DAMO bacteria could produce more oxygen for methane oxidation of type I aerobic methanotrophs and form a steady and efficient microbial community to remove nitrite [27].
Beyond any doubt, enrichment cultures are not only feasible substitutes of pure cultures to perform n-DAMO in wastewater treatment processes [23], but also the alternative samples for investigating physiological ecological and biochemical characteristics of the bacteria. AnSBR is an efficient reactor to enrich anaerobic bacteria due to its simple structure and unique setting phase during the operation, which microbes were not easily washed off. Thus, cornfield soil was inoculated into an anaerobic sequencing batch reactor (AnSBR) for the enrichment of n-DAMO bacteria in the present research. And then, microbial community structure of the acclimatized mixed culture was further revealed by the high-throughput sequencing to understand the role of n-DAMO bacteria in the cycle of carbon and nitrogen in anaerobic environment, which might give guidance for the application of n-DAMO bacteria on wastewater treatment.
2. Materials and Methods
2.1. Inoculum and Medium
The inoculum for the enrichment of n-DAMO bacteria was a soil sample collected from a cornfield after harvest. Geographic location of the sampling point and the specific characters of the sample were illustrated in Table 1. The collected sample was kept at 4°C for further use.
The medium according to the previous researches was used to enrich n-DAMO bacteria [13, 28]. The medium consisted of the following ingredients per liter of distilled water: 0.25 g KHCO3, 0.05 g KH2PO4, 0.3 g CaCl2·2H2O, 0.2 g MgSO4·7H2O, 0.069 g (1 mmol) NaNO2, 0.5 mL acidic trace element solution (100 mmol/L HCl) and 0.2 mL alkaline trace element solution. The acidic trace element solution (100 mmol/L HCl) contained (per liter): 2.085 g FeSO4·7H2O, 0.068 g ZnSO4·7H2O, 0.12 g CoCl2·6H2O, 0.5 g MnCl2·4H2O, 0.32 g CuSO4, 0.095 g NiCl2·6H2O and 0.014 g H3BO3. The alkaline trace element solution (10 mmol/L NaOH) contained (per liter): 0.067g SeO2, 0.050 g Na2WO4·2H2O and 0.242g Na2MoO4. The medium was flushed with nitrogen gas (N2) for 20 min to drive out oxygen (O2), and the pH adjusted to 7.0 – 7.5 by HCl (1 mol/L) and NaOH (1 mol/L).
2.2. Enrichment of n-DAMO Bacteria
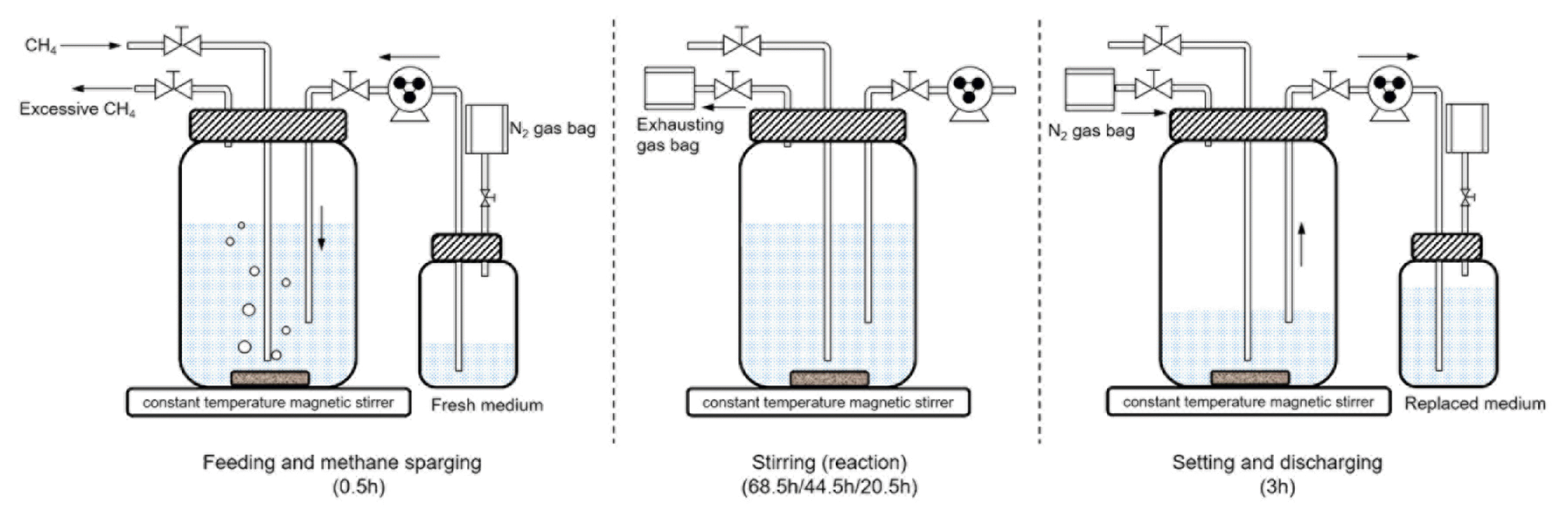
The reactor for n-DAMO bacteria enrichment was a 1.2 L glass bottle which was operated as an anaerobic sequencing batch reactor (AnSBR) (Fig. 1). The soil sample was sifted by a 0.25 mm mesh sieve after grinding and then inoculated into the reactor at a mixed liquor volatile suspended solids (MLVSS) of 6.0 g/L. Sealed with a rubber stopper, the reactor was loaded with 700 mL of the medium and then sparged with methane (99.999%) for 5 min acquiring about 350 mg CH4. The constructed reactor was put on a constant temperature magnetic stirrer (HDL apparatus, China) and incubated at 140 rpm and 30°C which is favorable for the growth of n-DAMO bacteria [14, 29]. The AnSBR was operated stage by stage with the operation cycle of 3, 2 or 1 d which was determined by the volumetric nitrite loading rate (VLR) of around 15.3, 23.0 and 46.0 mg/(L·d), respectively. By the end of each operation cycle, the AnSBR was stood for 3 h and then 400 mL of the supernatant was discharged. Fed with 400 mL of fresh medium within 0.5 hours and sparged with methane for 5 min, the renewed AnSBR was incubated again also at 30°C and 140 rpm. At the beginning and the end of each operation cycle, supernatant in the AnSBR was sampled for chemical analysis, respectively.
2.3. Analytical Methods
Nitrate (NO3−), NO2−, ammonium (NH4+), MLVSS and chemical oxygen demand (COD) were measured according to APHA standard methods [30]. Herein, NO3−, NO2−, and NH4+ were measured using colorimetric method. MLVSS was measured through gravimetric method. COD was quantified with the method of dichromate titration. NH4+, NO2− and NO3− in soil were extracted using 2 mol/L KCl (shaken for 1.5 h at 150 rpm) and then determined via APHA standard method. Soil temperature was measured using a temperature meter (Hach, USA). Soil pH was detected with a pH meter (Mettler Toledo, USA) after shaking soil water (1:5 w/v) suspension for 30 min [22]. A quantity of 200 mg soil sample was air-dried and sieved through a 0.25 mm mesh for analysis of total organic carbon (TOC). The TOC was measured by an element analyzer (Vario ELcube, Elementar, Germany).
The volume fraction of CH4 and CO2 in biogas was measured on a gas chromatograph (SP-6800A, China) equipped with a 2 m stainless column (packed with Porapak Q (100 to 180 mesh)) and a thermal conductivity detector (TCD) [31]. Temperature of the oven, injection port, and detector were all set at the same 80°C. N2 was used as the carrier gas at a flow rate of 40 mL/min. The injection volume was 0.5 mL.
2.4. High-throughput Sequencing
To understand the microbial community structure of the mixed culture, the suspended sludge in the AnSBR was sampled by the end of the enrichment process (the 232nd day) and then delegated to Sangon Biotech Co., Ltd (Shanghai, China) for high-throughput sequencing. DNA of the sample was isolated with E.Z.N.A Soil DNA Kit (OMEGA, USA) according to the manufacture’s instruction. Bacterial 16S rDNA for the V3–V4 regions were amplified with primer pairs 341f (5′-CCTACGGGAGGCAGCAG-3′) and 805r (5′-GACTACHVGGGTATCTAATCC-3′) [32]. Composition of V3–V4 regions in the PCR products was sequenced on the Illumina Miseq sequencing platform. Raw sequences were filtered to remove low-quality reads and clustered into Operational Taxonomic Units (OTUs) at 97% of sequence similarity using QIIME v1.8.0 (USA) according to the NCBI taxonomy database. The community diversity and richness were calculated using Mothur 1.30.1. Sequence clusters were created using NCBI Blast+ 2.28 (USA). Indices of Chaos, ACE, Shannon, Simpson and Coverage were all calculated by the online website software (http://www.mothur.org/).
3. Results and Discussion
3.1. Enrichment Process of n-DAMO Bacteria
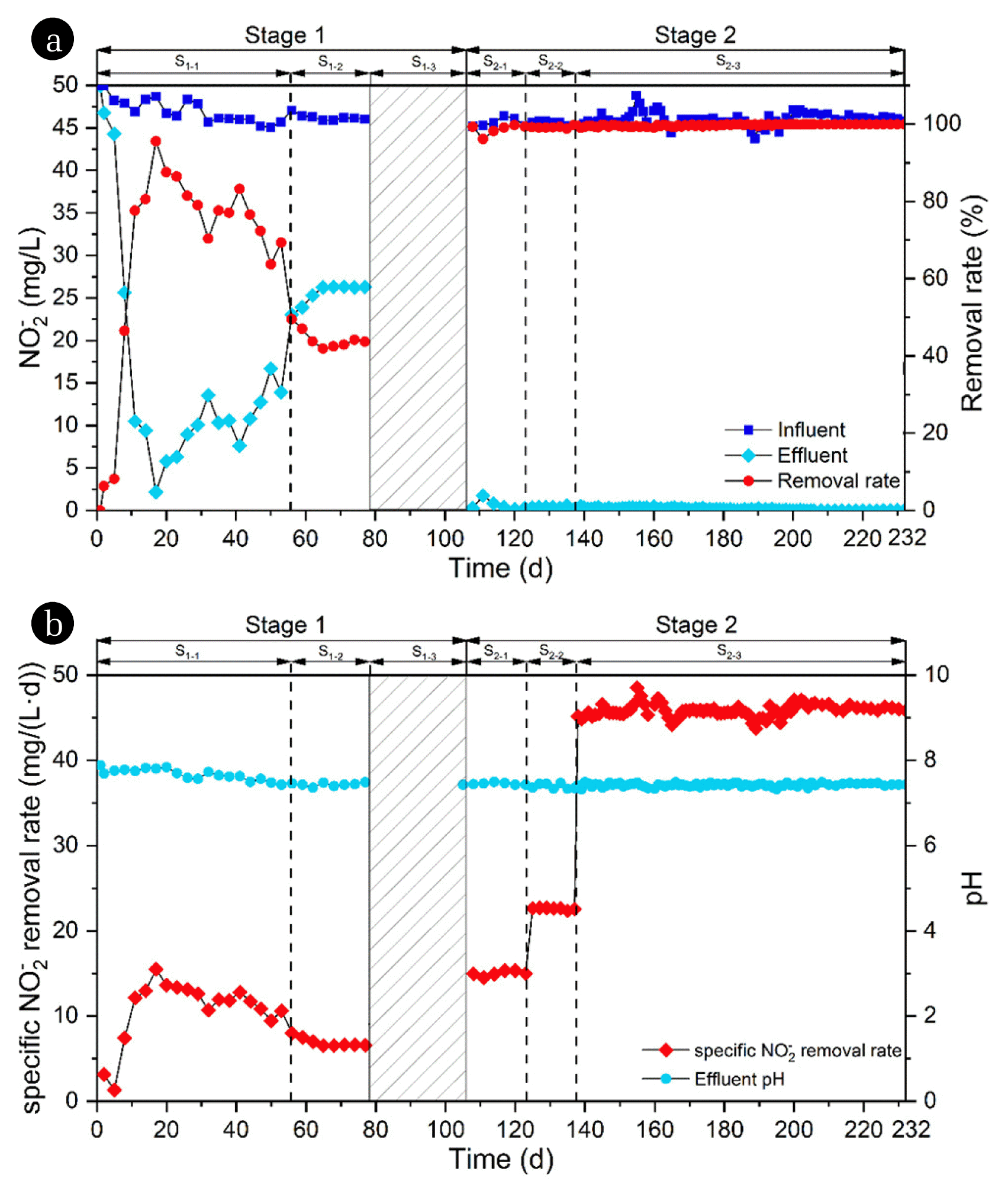
Inoculated with 6.0 g MLVSS/L of the cornfield soil, enrichment of n-DAMO bacteria was conducted in the AnSBR for 232 d. All through the enrichment process, the NO2− in feed was kept at about 46.0 mg/L to avoid the possible toxicity to bacteria [33]. As illustrated in Fig. 2, the enrichment process was divided into two stages, with the first 104 d and the last 128 d as Stage 1 (S1) and Stage 2 (S2), respectively. Considering the slow growth and poor adaptability of n-DAMO bacteria to a strange environment, the AnSBR was started up at an operational cycle of 3 d till to the 53rd day, which was marked as the first phase (S1-1) of S1 by the VLR of NO2− as low as 15.3 mg/(L·d). As shown in Fig. 2(a), NO2− removal in the reactor was rapidly increased to 95.6% within the first 17 d, with 2.17 mg/L of NO2− remaining in the effluent. However, the removal efficiency of NO2− decreased gradually within the next 38 d. By the end of S1-1 (the 55th day), the residual NO2− in effluent averaged 13.8 mg/L with a removal rate as low as 69.3% in the reactor. To encourage the growth of n-DAMO bacteria, the VLR of NO2− was increased to 23.0 mg/(L·d) by shortening the operational cycle to 2 d in the second phase (S1-2) of S1. However, NO2− removal in the reactor was further decreased at the NO2− VLR of 23.0 mg/(L·d). The NO2− removal during the steady state (62nd to 77th days) of S1-2 averaged 43.2% (26.3 mg/L in effluent), suggesting the metabolic activity of n-DAMO bacteria was persistently inhibited in the AnSBR.
In the present research work, the inoculum was the soil sampled from a cornfield after harvest, which was rich in biodegradable organics (Table 1). It was thought that the degradation of organic carbon sources had stimulated the growth of chemoheterotrophic microorganisms, resulting in the restriction of n-DAMO bacteria in the AnSBR [34]. It is known that hydrolysis of organics is the rate-limiting step of anaerobic digestion [35]. With few hydrolysates of organics, the growth of chemoheterotrophic microorganisms should be badly restricted in the first 17 d of S1, which would provide an opportunity for the multiplication of autotrophic n-DAMO bacteria (Fig. 2(a)). Along with the extension of culturing time, more hydrolysates were released from the organics, resulting in the flourishing of chemoheterotrophic bacteria in the AnSBR. With the slow growth and metabolism, the autotrophic n-DAMO bacteria must have been obviously restricted since the 18th day in S1 [36]. To avoid the negative effect of chemoheterotrophic bacteria on the enrichment of n-DAMO bacteria, the AnSBR went through a 27-day idle period, the third phase (S1-3) of S1, to exhaust the available organic carbon sources.
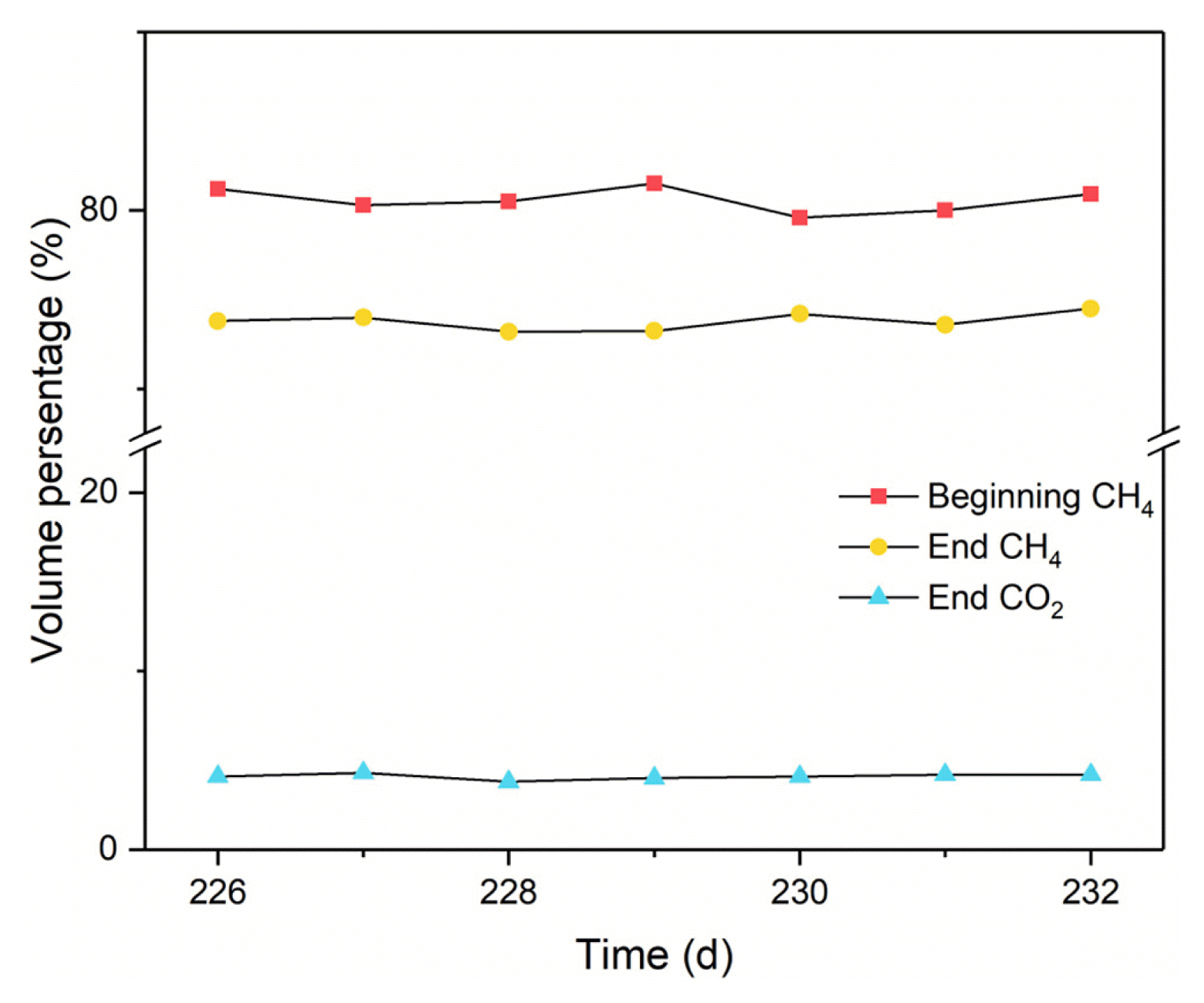
After the idle period (S1-3), the AnSBR was restarted at the NO2− VLR of 15.3 mg/(L·d) with the operation cycle of 3 d, which was marked as the first phase (S2-1) of S2. At the beginning of S2-1, the COD concentration in the reactor decreased under the detection limit. As expected, the bad NO2− removal in S1 was restored in S2. As shown in Fig. 2(a), NO2− in feed was almost exhausted all through the 128 d in S2, although set NO2− VLR had been increased from 15.3 to 23.0 and then to 46.0 mg/(L·d) with the operation cycle of 3, 2 and 1 d in the S2-1, second phase (S2-2) and third phase (S2-3), respectively. CH4 consumption and CO2 production were found along with the excellent removal of NO2−. As shown in Fig. 3, the average volume fraction of CH4 in the gaseous phase was decreased from the initial 80.6% to 73.8% by the end of the operational cycles, along with 4.1% CO2 detected, in S2-3. The continuous CH4 consumption and CO2 production along with the excellent removal of NO2− indicated that n-DAMO bacteria had been enriched stage by stage [11, 14]. Within the last phase S2-3, the actual specific NO2− removal rate in the AnSBR averaged 46.16 mg/(L·d) with a constant pH of about 7.4 (Fig. 2(b)) that was attributed to the buffers of KHCO3 and KH2PO4 in the medium. By the end of S2-3, MLVSS in the AnSBR was detected to be 6.87 g/L, i.e. the specific NO2− removal rate of the acclimatized mixed culture was about 6.72 mg/(g MLVSS·d).
3.2. Community Structure of the Acclimatized Mixed Culture
On the 232nd day, the mixed culture in the AnSBR was sampled for high-throughput sequencing to reveal the microbial community structure and functional bacterial diversity. With the good coverage of 0.97, 35942 clean reads were obtained and clustered into 1404 OTUs at 97% of sequence similarity. Indices Shannon, Simpson, Chao1 and ACE of the mixed culture were 4.24, 0.04, 4974.12 and 9259.12, respectively. The results suggested that the species of the enrichment were diverse [37].
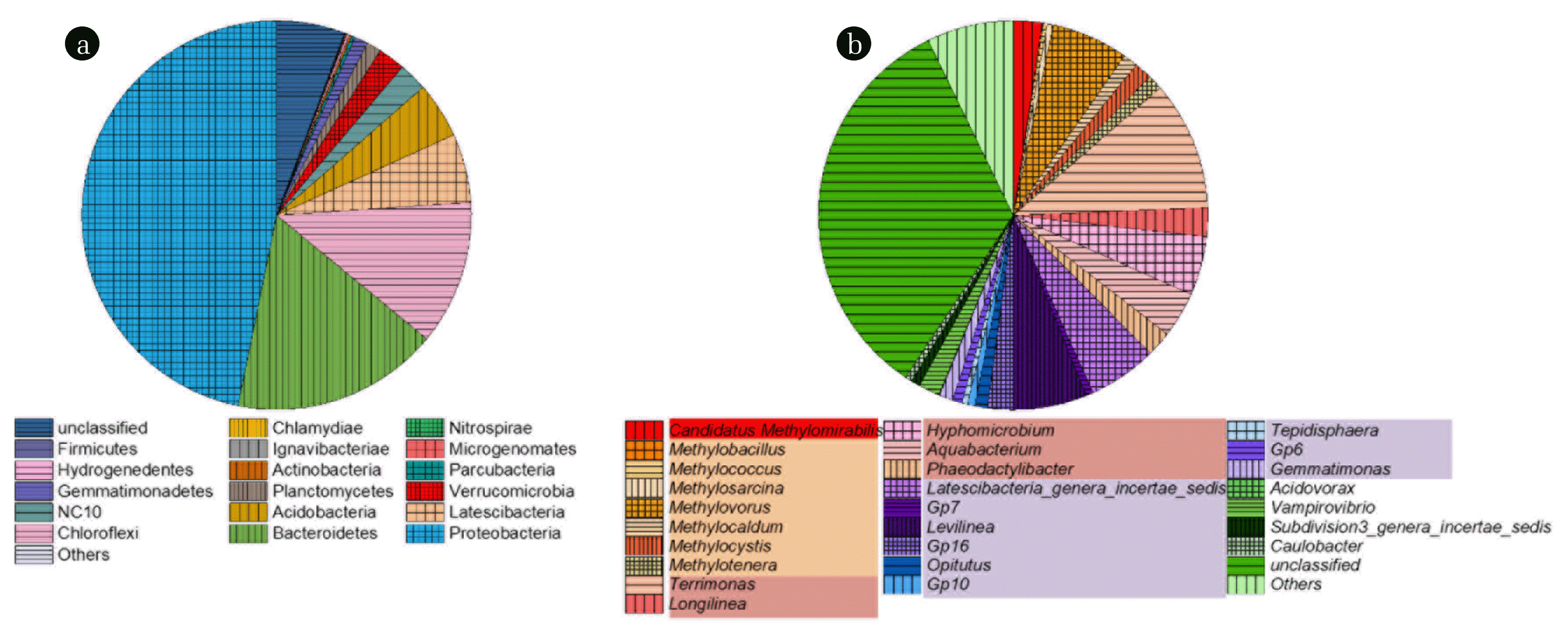
To reveal the removal pathways of NO2− in the mixed culture, microbial community structure on phylum and genus level was further analyzed. The phylum assignment (Fig. 4(a)) showed that Proteobacteria, Bacteroidetes and Chloroflexi were the most abundant three phyla in the culture, with a relative abundance as high as 46.81%, 17.2% and 12.03%, respectively. Most denitrifying bacteria are classified into the three phyla [38]. Latescibacteria and Acidobacteria were also rich in the mixed culture with a relative abundance of 5.73% and 4.65%, respectively. Bacteria affiliated to Latescibacteria mainly exist in soil and function in degrading plant polysaccharide [39]. The bacteria in Acidobacteria can adapt to a low-nutrient environment [40]. As one of the phyla in which methanotrophs were cataloged, the relative abundance of Verrucomicrobia reached 2.27%. Phylum NC10 has collected all of the identified n-DAMO bacteria [17], and the relative abundance reached 2.37% in the mixed culture.
As illustrated in Fig. 4(b), the unclassified bacteria accounted for 33.48% in the mixed culture, most of which were indicated as the bacteria in phylum Proteobacteria. Besides the unclassified bacteria, Terrimonas, Methylovorus, Levilinea, Latescibacteria_genera_ incertae_sedis and Hyphomicrobium were the most abundant genera, with a relative abundance of 10.75%, 6.36%, 6.12%, 5.73% and 4.9%, respectively. Although Terrimonas is classified as an aerobic heterotrophic genus, some bacteria in the genus have been detected in anaerobic conditions [41]. Methylovorus has been described as a group of restricted facultative methylotrophic bacteria, characterized by the assimilation of methanol via ribulose monophosphate pathway and the presence of glutamate cycle enzymes for ammonia assimilation [42]. Levilinea has been indicated as acidogenic fermenters which can degrade various carbohydrates to volatile acids [43]. Chemoheterotrophic bacteria Latescibacteria_genera_incertae_sedis, mainly distributed in soil [39], was also found in the mixed culture. Hyphomicrobium has been reported as denitrifying bacteria which can convert NO2− to N2 using methanol as a carbon source in anaerobic condition [44]. As the only reported genus performing n-DAMO, Candidatus Methylomirabilis accounted for 2.37% in the mixed culture. Some other methanotrophs were also found in the mixed culture, such as Methylocystis, Methylocaldum, Methylophilus, Methylosarcina and Methylococcus. All of the bacteria in the five genera have been identified as aerobic bacteria which convert methane to one-carbon substances such as methanol [45]. It was concluded that both aerobic and anaerobic methane-oxidizing bacteria coexisted in the mixed culture, which agreed with the earlier research [46]. The diversity of species and the complex microbial community suggested that a complete food web had been caused by the supplied CH4 and NO2− as essential nutrients.
3.3. Biochemical Characteristics and Nitrite Removal Approaches in the Mixed Culture
The biodegradable organics in the soil sample (Table 1) must play a role as the electronic donors to heterotrophic denitrification of NO2−, which resulted in the rapid increase of NO2− removal in the AnSBR within the first 17 d. The gradually decreased NO2− removal (within the days from the 18th to 80th day) indicated the exhaustion of biodegradable organics by the end of S1-2. Although the hydrolysates of organics in the soil could also contribute to the heterotrophic denitrification, the availability should have been severely restricted due to the refractory to biodegradation [47]. Thus, n-DAMO was identified as a dominant approach to the good NO2− removal in S2-3. However, the good NO2− removal should be challenged by the relative abundance of n-DAMO bacteria (Candidatus Methylomirabilis) as low as 2.37% in the mixed culture.
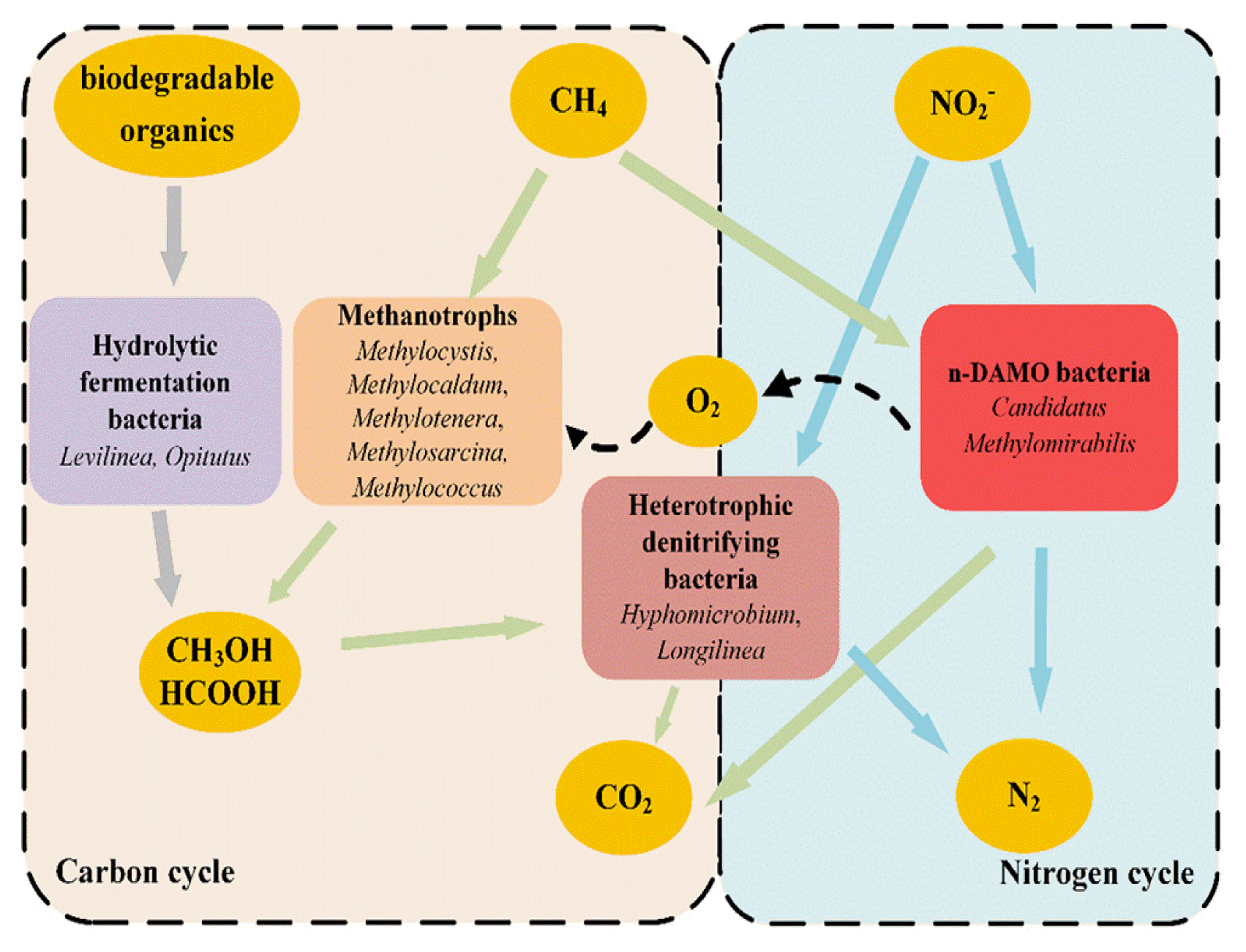
As shown in Fig. 4(b), the community structure of the mixed culture was complex with the coexistence of aerobic heterotrophic bacteria (Terrimonas), facultative anaerobic bacteria (Methylovorus, Aquabacterium), carbohydrate fermenting bacteria (Levilinea), chemoheterotrophic bacteria (Latescibacteria), denitrifying bacteria (Hyphomicrobium) and n-DAMO bacteria (Candidatus Methylomirabilis). The diversity of species and biochemical characteristics suggested a complex food web was established in the AnSBR. Though it was difficult to reveal the metabolic network in the mixed culture in detail, the following cooperation between the identified dominant genera could be identified as illustrated in Fig. 5: 1) the supplied NO2− was reduced to N2 by Candidatus Methylomirabilis via n-DAMO with the supplied CH4 as electronic donors, during which O2 would be produced (Eq. (1), Eq. (2), and Eq. (3)) [48, 49]; 2) the produced O2 would further be absorbed and utilized by methanotrophs (Methylocystis, Methylocaldum, Methylotenera, Methylosarcina and Methylococcus) for the biosynthesis of biomass, methanol and formic acid [27, 45]; 3) the synthesized methanol and formic acid by the methanotrophs, as well as the organic acids formed by hydrolytic fermentation bacteria (Levilinea and Opitutus) [43], all served as the electronic donors for the reduction of NO2− by denitrifying bacteria (Hyphomicrobium and Longilinea) with CO2 and N2 as the end products [50].
Above all, the nitrite removal approaches had been revealed in the mixed culture of n-DAMO bacteria, based on the microbial species and supplied nutrients, CH4 and NO2−. Though n-DAMO had been identified as the dominant approach to NO2− removal, assimilation and heterotrophic denitrification had also contributed to the NO2− removal in the AnSBR. The diversity of species and biochemical characteristics suggested that the mixed culture had a high availability for nitrogen removal in wastewater treatment plants.
4. Conclusions
The mixed culture that contained n-DAMO bacteria was acquired in an AnSBR after 232-day cultivation inoculated from cornfield soil. The relative abundance of n-DAMO bacteria reached 2.37% in the enrichment. The specific NO2− removal rate of the mixed culture was as high as 6.72 mg/(g MLVSS·d) with the NO2− load removal of about 46.16 mg/(L·d) in the AnSBR. The reactor showed a complex community structure with the coexistence of n-DAMO bacteria, methanotrophs, heterotrophic denitrifying bacteria and hydrolytic fermentation bacteria. By analyzing the biochemical characteristics and microbial community structure, we found that n-DAMO bacteria, Candidatus Methylomirabilis, played an important role in combining nitrogen cycle and carbon cycle. The present work opens a direction for the enrichment of n-DAMO bacteria and proposed the possible application of n-DAMO bacteria in nitrite removal of the wastewater treatment plant.